Difference between revisions of "Team:Leiden/Demonstrate"
| Line 1: | Line 1: | ||
| − | + | <html lang="en"> | |
| − | <html> | + | |
| + | <head> | ||
| + | <meta charset="UTF-8"> | ||
| + | <meta name="viewport" content="width=device-width, initial-scale=1.0"> | ||
| + | <meta http-equiv="X-UA-Compatible" content="ie=edge"> | ||
| + | <meta name="author" content="Chiel van Amstel"> | ||
| + | <link rel="stylesheet" href="https://2018.igem.org/Team:Leiden/css/main_css?action=raw&ctype=text/css" type='text/css'> | ||
| + | <link rel="stylesheet" href="https://2018.igem.org/Team:Leiden/css/fontawesome-all_min_css?action=raw&ctype=text/css" type="text/css"> | ||
| + | <link rel="stylesheet" href="https://2018.igem.org/Team:Leiden/css/navbarfooter_css?action=raw&ctype=text/css" type="text/css"> | ||
| + | </head> | ||
| + | <body> | ||
| + | <div class="no-js" id="loader-wrapper"> | ||
| + | <div id="loader"></div> | ||
| + | <div id="BestOnLargeScreens">Although the mobile version has the same content, this wiki is best viewed on larger screens </div> | ||
| + | </div> | ||
| + | <script type="text/javascript"> | ||
| + | document.querySelector(".no-js").classList.remove('no-js'); | ||
| + | var Timestamp; | ||
| + | // var Timestamp = Date.now(); | ||
| + | </script> | ||
| + | <nav id="header_html" data-source="https://2018.igem.org/Team:Leiden/navbar_html?action=raw"></nav> | ||
| − | |||
| + | <div class="mainContent"> | ||
| + | <div class="container-text"> | ||
| + | <h1 class="SectionTitle">Demonstration</h1> | ||
| + | Antibiotic discovery is in sore need of innovation. Current antibiotic screening focuses on detecting lethal antimicrobial compounds. This leads to many non-lethal - but stressful - compounds being missed by conventional testing. Therefore, we | ||
| + | propose a novel method for antibiotic screening that allows stressful compounds to be detected as well. Compounds that cause stress could be used in synergistic combination therapies that will delay the onset of antibiotic resistance. | ||
| + | Additionally, our method allows detection of non-lethal concentrations of lethal compounds. With our system we hope to drive the antibiotic discovery field forward by enabling easy detection of bacterial cell stress. To this end, we have | ||
| + | developed two frameworks for the detection of stressful compounds to be used in the lab by antibiotic researchers in academics and industry. | ||
| + | </div> | ||
| + | <section class="popout chapter"> | ||
| + | <div class="collapsible"> Bacterial cell stress visualised for the naked eye </div> | ||
| + | <div class="collapsibleContent"> | ||
| + | <h2 class="SubSectionTitle"></h2> | ||
| + | <p>Bacterial overlays are typically performed using <i>Streptomyces</i> or other antibiotic producing bacterial colonies. To perform an overlay experiment, an agar plate is dotted with colonies of a (pathogenic) organism of interest. These | ||
| + | antibiotic producing colonies secrete compounds that are damaging to the surrounding cells, creating a so called ‘death halo’ around the colony. This halo indicates a bactericidal activity of the producing colony. Afterwards the secreted | ||
| + | compounds that caused the halo are isolated and analysed for their structure and possible mechanism of action. Many of our current-day antibiotics have been discovered using these techniques. However, this process can be very time | ||
| + | consuming. For this reason, we have created a reporter strain in <i>E. coli </i> DH5α using chromoproteins coupled to stress activated promoters. When these <i>E. coli</i> cells are applied to an agar plate with an antibiotics | ||
| + | producing colony, they | ||
| + | will produce a color when being stressed. This not only indicates a mechanism of action for these antibiotics early in the discovery process - in the form of a specific chromoprotein colour produced by a specific stress promoter - but | ||
| + | also allows us to detect stressful but not necessarily deadly compounds and non-lethal concentrations of lethal compounds.<br><br> For demonstration purposes, we have replicated these bacterial overlays using a traditional disk diffusion | ||
| + | assay. Disk diffusion assays encompass antibiotic disks being placed on test organism saturated agar plates. These disks release the antibiotic slowly creating death halos around the disk (Figure 1). Thus, they are very similar in function | ||
| + | to bacterial overlays and provide an excellent replacement for demonstration purposes. </p> | ||
| + | <figure class="resultfig horizontal"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/1/1f/T--Leiden--diskmin.jpg"></img> | ||
| + | <figcaption><b>Figure 1.Disk diffusion on an <i>E. coli</i> saturated Mueller-Hinton agar plate with four nalidixic acid disks</b> </figcaption> | ||
| + | </figure> | ||
| + | <p> | ||
| + | In order to test our colourful bacterial strains, we saturated a Mueller-Hinton agar plate with pSoxS-AmilCP (<a href="http://parts.igem.org/Part:BBa_K2610033" class="colouredlink" target="_blank">BBa_K2610033</a>) transformed <i>E. coli</i> | ||
| + | DH5α - one of our stress response elements, capable of detecting DNA damage. A disk containing nalidixic acid, an antibiotic which selectively and reversibly blocks DNA replication, was placed on the pSoxS-AmilCP (<a href="http://parts.igem.org/Part:BBa_K2610033" | ||
| + | class="colouredlink" target="_blank">BBa_K2610033</a>) expressing bacteria. Figure 2 shows an example of such a disk diffusion. Around the disk - visible in white - the death halo is apparent, additionally, at the edge of the halo the | ||
| + | <i>E. coli</i> bacteria are coloured blueish grey. This shows that as a result of the upregulation of pSoxS in these cells, AmilCP is being produced. This is indicative of a stressful process occurring in these cells. Hereby, we have shown | ||
| + | that | ||
| + | non-lethal concentrations of nalidixic acid can be detected using disk diffusion with our chromoproteins stress reporter BioBricks. | ||
| + | </p> | ||
| + | <figure class="resultfig horizontal"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/2/2c/T--Leiden--Model3diskOverlay.jpg"></img> | ||
| + | <figcaption><b>Figure 2. Disk diffusion assay performed with <i>E. coli</i> DH5α expressing pSoxS-AmilCP (<a href="http://parts.igem.org/Part:BBa_K2610033" class="colouredlink" target="_blank">BBa_K2610033</a>), showing a typical | ||
| + | death | ||
| + | halo but also with a stress induced blue/greyish chromoprotein halo</b> </figcaption> | ||
| + | </figure> | ||
| + | </div> | ||
| + | </section> | ||
| − | <div class=" | + | <section class="popout chapter"> |
| − | < | + | <div class="collapsible">GFP based stress detection system responds selectively to antibiotics</div> |
| − | < | + | <div class="collapsibleContent"> |
| + | <h2 class="SubSectionTitle"></h2> | ||
| + | <p>Our system is designed to selectively respond to antibiotics based on their cellular targets, while not responding to other antibiotics targeting other cellular targets. This allows for early estimations about the mechanism of | ||
| + | action of candidate compounds, thereby shortening drug discovery pipelines. Some of the BioBricks we demonstrated this objective with, are pCspA-GFP (<a href="http://parts.igem.org/Part:BBa_K2610037" class="colouredlink" target="_blank">BBa_K2610037</a>), | ||
| + | pKatG-GFP (<a href="http://parts.igem.org/Part:BBa_K2610018" class="colouredlink" target="_blank">BBa_K2610018</a>) and pSoxS-GFP (<a href="http://parts.igem.org/Part:BBa_K2610031" class="colouredlink" target="_blank">BBa_K2610031</a>), | ||
| + | using flow cytometry. Cells were stressed by a repertoire of known antibiotics at non-lethal concentrations, in order to obtain their unique response profiles, such as can be seen in figure 3A, B and C for pCspA, pSoxS and pKatG | ||
| + | respectively. Promoter CspA is suppressed by nearly all antibiotics, but is activated specifically by nalidixic acid. In contrast to pCspA, promoter SoxS is not only activated by nalidixic acid but also by zeocin (both associated with DNA | ||
| + | damage). Conversely, promoter KatG | ||
| + | is activated only by hydrogen peroxide. Therefore, we have shown that our stress activated promoters react only to specific stressors. This is vitally important for our screening system because it allows for differentiation between modes | ||
| + | of actions of candidate compounds. | ||
| + | </p> | ||
| + | <figure class="resultfig"> | ||
| + | <div class="FigFlexbox"> | ||
| + | <div class="flexfig"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/5/58/T--Leiden--DemonstrateFig3A.png"></div> | ||
| + | <div class="flexfig"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/5/58/T--Leiden--DemonstrateFig3B.png"></div> | ||
| + | <div class="flexfig"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/5/55/T--Leiden--DemonstrateFig3C.png"></div> | ||
| + | </div> | ||
| + | <figcaption><b>Figure 3 Specificity of stress response</b> These results show distinctive responses for each of the promoters. | ||
| + | <!-- caption? --> | ||
| + | </figcaption> | ||
| + | </figure> | ||
| − | < | + | <!-- Figure 3ABC hier--> |
| − | + | ||
| − | + | ||
| − | <p> | + | </div> |
| − | + | </section> | |
| − | </p> | + | <section class="popout chapter"> |
| + | <div class="collapsible">GFP based stress detection reveals mode of action and enhances antibiotics. </div> | ||
| + | <div class="collapsibleContent"> | ||
| + | <h2 class="SubSectionTitle"></h2> | ||
| + | <p>Combination therapies have been proven to suppress the development of resistance in cancer and HIV treatment<sup>[1][2]</sup>. Moreover, synergistic combination therapies for fighting infections are already being used in the clinic in the | ||
| + | form of | ||
| + | amoxicillin/clavulanic acid. During our research we have proven that a non-lethal stressful compound verified by our system could be used to enhance the effect of antibiotics. We screened a number of natural and chemically engineered | ||
| + | compounds for their ability to stress bacterial cells, in this case <i>E. coli</i> DH5α. Firstly, compounds that were lethal were eliminated from the screen to ensure results from our system were in response to purely stressful | ||
| + | compounds (Figure 4). | ||
| + | Consequently, we continued with a select number of compounds due to time restraints. The following compounds were selected through literature research: ginger, ascorbic acid, cinnamon and cannabis<sup>[3-8]</sup>. We tested the ability of | ||
| + | these compounds | ||
| + | to activate or suppress various stress-related promoters fused to GFP using flow cytometry. </p> | ||
| + | <figure class="resultfig horizontal"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/7/72/T--Leiden--demonstratetrechter.png"></img> | ||
| + | <figcaption class="center"><b>Figure 4. Experimental design of our system demonstration</b> </figcaption> | ||
| + | </figure> | ||
| + | <p>The clpB promoter (<a href="http://parts.igem.org/Part:BBa_K2610020" class="colouredlink" target="_blank">BBa_K2610020</a>), a gene involved in the cellular stress response to aggregated proteins, responded to ascorbic acid, more commonly | ||
| + | known as vitamin C (Figure 5). This is an indication that this compound is involved in protein aggregation. Previous research has shown an involvement of ascorbic acid in the inactivation of proteins<sup>[9][10]</sup>. This suggests our | ||
| + | screening | ||
| + | system can indicate a possible mechanism of action for stressful compounds. </p> | ||
| + | <figure class="resultfig wide"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/9/98/T--Leiden--VITCclpBresult.png"></img> | ||
| + | <figcaption><b>Figure 5. pClpB responds to ascorbic acid with an increased fluorescent response</b> </figcaption> | ||
| + | </figure> | ||
| + | <p>Ascorbic acid not only emerged as a stressful substance but we also demonstrated that this compound could lower the minimum inhibitory concentration (MIC) of ampicillin (Figure 6A), chloramphenicol (Figure 6B) and nalidixic acid (Figure | ||
| + | 6C). | ||
| + | This proves our system is able to detect stressful compounds which can be used in antibiotic combination therapies.</p> | ||
| + | <figure class="resultfig"> | ||
| + | <div class="FigFlexbox"> | ||
| + | <div class="flexfig"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/4/43/T--Leiden--DemonstrateFig6A.png"></div> | ||
| + | <div class="flexfig"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/5/56/T--Leiden--DemonstrateFig6B.png"></div> | ||
| + | <div class="flexfig"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/8/8c/T--Leiden--DemonstrateFig6C.png"></div> | ||
| + | </div> | ||
| + | <figcaption><b>Figure 6. Addition of ascorbic acid to ampicillin treatment (A), chloramphenicol treatment (B) and nalidixic acid treatment (C) causes a reduction in minimum inhibitory concentration.</b> </figcaption> | ||
| + | </figure> | ||
| − | </div> | + | <div class="smaller refs"> |
| + | <h3>References</h3> | ||
| + | <p id="Ref1">[1]: Yardley, D. A. (2013). Drug resistance and the role of combination chemotherapy in improving patient outcomes. International journal of breast cancer, 2013. | ||
| + | </p> | ||
| + | <p id="Ref2">[2]: Katlama, C., Ingrand, D., Loveday, C., Clumeck, N., Mallolas, J., Staszewski, S., ... & McDade, H. (1996). Safety and efficacy of lamivudine-zidovudine combination therapy in antiretroviral-naive patients: a randomized | ||
| + | controlled comparison with zidovudine monotherapy. Jama, 276(2), 118-125. | ||
| + | </p> | ||
| + | <p id="Ref3">[3]: Tajkarimi, M., & Ibrahim, S. A. (2011). Antimicrobial activity of ascorbic acid alone or in combination with lactic acid on Escherichia coli O157:H7 in laboratory medium and carrot juice. Food Control, 22(6), 801–804. | ||
| + | https://doi.org/10.1016/j.foodcont.2010.11.030 | ||
| + | </p> | ||
| + | <p id="Ref4">[4]: Mathew, S., Verghese, R., & David, A. (2017). Antimicrobial activity of Vitamin C demonstrated on uropathogenic Escherichia coli and Klebsiella pneumoniae. Journal of Current Research in Scientific Medicine, 3(2), 88. | ||
| + | https://doi.org/10.4103/jcrsm.jcrsm_35_17 | ||
| + | </p> | ||
| + | <p id="Ref5">[5]: Ali, E. M. M., Almagboul, A. Z. I., Khogali, S. M. E., & Gergeir, U. M. A. (2012). Antimicrobial Activity of Cannabis sativa L. Chinese Medicine, 03(01), 61–64. https://doi.org/10.4236/cm.2012.31010 | ||
| + | </p> | ||
| + | <p id="Ref6">[6]: Chakraborty, S., Afaq, N., Singh, N., & Majumdar, S. (2018). Antimicrobial activity of Cannabis sativa, Thuja orientalis and Psidium guajava leaf extracts against methicillin-resistant Staphylococcus aureus. Journal of | ||
| + | Integrative Medicine, 16(5), 350–357. https://doi.org/10.1016/j.joim.2018.07.005 | ||
| + | </p> | ||
| + | <p id="Ref7">[7]: Patel, R. V., Thaker, V. T., & Patel, V. (2011). Antimicrobial activity of ginger and honey on isolates of extracted carious teeth during orthodontic treatment. Asian Pacific Journal of Tropical Biomedicine, 1(1), | ||
| + | 867–871. https://doi.org/10.1016/s2221-1691(11)60124-x | ||
| + | </p> | ||
| + | <p id="Ref8">[8]: Gupta, S., & Ravishankar, S. (2005). A Comparison of the Antimicrobial Activity of Garlic, Ginger, Carrot, and Turmeric Pastes Against Escherichia coli O157:H7 in Laboratory Buffer and Ground Beef. Foodborne Pathogens and | ||
| + | Disease, 2(4), 330–340. https://doi.org/10.1089/fpd.2005.2.330 | ||
| + | </p> | ||
| + | <p id="Ref9">[9]: Szarka, A., & Lőrincz, T. (2014). The role of ascorbate in protein folding. Protoplasma, 251(3), 489-497. | ||
| + | </p> | ||
| + | <p id="Ref10">[10]: Stadtman, E. R. (1991). Ascorbic acid and oxidative inactivation of proteins. The American journal of clinical nutrition, 54(6), 1125S-1128S. | ||
| + | </p> | ||
| + | </div> | ||
| + | </div> | ||
| + | </section> | ||
| + | <!-- end of main content here --> | ||
| + | </div> | ||
| + | <div id="sidenav" class="farRight"> | ||
| + | <span id="ClickToCollapseSidenav"><i class="fas fa-angle-left"></i></span> | ||
| + | </div> | ||
| + | <div class="UnderPageNav"> | ||
| + | <a href="https://2018.igem.org/Team:Leiden/Results" class="NavBlock"> | ||
| + | <div class="NavBlockIcon"><i class="icon Results"></i></div> | ||
| + | <div class="NavBlockText"><span>Results</span>Click here to read about all our results | ||
| + | </div> | ||
| + | </a> | ||
| + | <a href="https://2018.igem.org/Team:Leiden/Applied_Design" class="NavBlock"> | ||
| + | <div class="NavBlockIcon"><i class="icon Applied-design"></i></div> | ||
| + | <div class="NavBlockText"><span>Product Design</span>Click here to read how our demonstrated system lead to a product design | ||
| + | </div> | ||
| + | </a> | ||
| + | <a href="https://2018.igem.org/Team:Leiden/Entrepreneurship" class="NavBlock"> | ||
| + | <div class="NavBlockIcon"><i class="icon Entrepreneurship"></i></div> | ||
| + | <div class="NavBlockText"><span>Entrepreneurship</span>Click here to read how we plan to bring our demonstrated product to the market | ||
| + | </div> | ||
| + | </a> | ||
| + | </div> | ||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | <footer id="footer_html" data-source="https://2018.igem.org/Team:Leiden/Footer?action=raw" style="color:var(--blackish)"></footer> | ||
| + | |||
| + | |||
| + | <script src="https://2018.igem.org/Team:Leiden/javascript/jquery-3_3_1_min_js?action=raw&ctype=text/javascript" type="text/javascript"></script> | ||
| + | <script src="https://2018.igem.org/Team:Leiden/javascript/removeBullshitClasses_js?action=raw&ctype=text/javascript" charset="utf-8"></script> | ||
| + | <script src="https://2018.igem.org/Team:Leiden/javascript/main_js?action=raw&ctype=text/javascript" charset="utf-8" type="text/javascript"></script> | ||
| + | <script type="text/javascript"> | ||
| + | $(document).ready(function() { | ||
| + | window.include('#footer_html'); | ||
| + | window.include('#header_html'); | ||
| + | }); | ||
| + | </script> | ||
| + | </body> | ||
</html> | </html> | ||
Revision as of 15:45, 15 October 2018
Demonstration
Antibiotic discovery is in sore need of innovation. Current antibiotic screening focuses on detecting lethal antimicrobial compounds. This leads to many non-lethal - but stressful - compounds being missed by conventional testing. Therefore, we propose a novel method for antibiotic screening that allows stressful compounds to be detected as well. Compounds that cause stress could be used in synergistic combination therapies that will delay the onset of antibiotic resistance. Additionally, our method allows detection of non-lethal concentrations of lethal compounds. With our system we hope to drive the antibiotic discovery field forward by enabling easy detection of bacterial cell stress. To this end, we have developed two frameworks for the detection of stressful compounds to be used in the lab by antibiotic researchers in academics and industry.Bacterial overlays are typically performed using Streptomyces or other antibiotic producing bacterial colonies. To perform an overlay experiment, an agar plate is dotted with colonies of a (pathogenic) organism of interest. These
antibiotic producing colonies secrete compounds that are damaging to the surrounding cells, creating a so called ‘death halo’ around the colony. This halo indicates a bactericidal activity of the producing colony. Afterwards the secreted
compounds that caused the halo are isolated and analysed for their structure and possible mechanism of action. Many of our current-day antibiotics have been discovered using these techniques. However, this process can be very time
consuming. For this reason, we have created a reporter strain in E. coli DH5α using chromoproteins coupled to stress activated promoters. When these E. coli cells are applied to an agar plate with an antibiotics
producing colony, they
will produce a color when being stressed. This not only indicates a mechanism of action for these antibiotics early in the discovery process - in the form of a specific chromoprotein colour produced by a specific stress promoter - but
also allows us to detect stressful but not necessarily deadly compounds and non-lethal concentrations of lethal compounds.
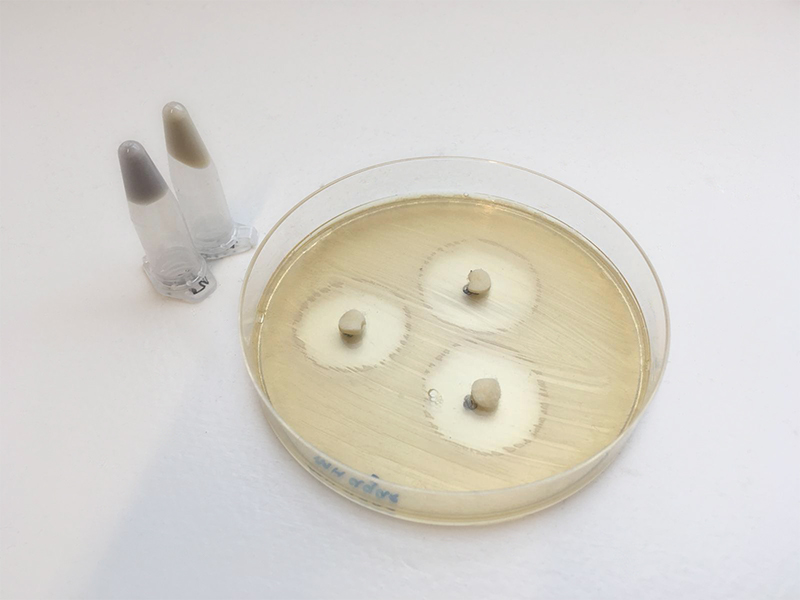
For demonstration purposes, we have replicated these bacterial overlays using a traditional disk diffusion
assay. Disk diffusion assays encompass antibiotic disks being placed on test organism saturated agar plates. These disks release the antibiotic slowly creating death halos around the disk (Figure 1). Thus, they are very similar in function
to bacterial overlays and provide an excellent replacement for demonstration purposes.

In order to test our colourful bacterial strains, we saturated a Mueller-Hinton agar plate with pSoxS-AmilCP (BBa_K2610033) transformed E. coli DH5α - one of our stress response elements, capable of detecting DNA damage. A disk containing nalidixic acid, an antibiotic which selectively and reversibly blocks DNA replication, was placed on the pSoxS-AmilCP (BBa_K2610033) expressing bacteria. Figure 2 shows an example of such a disk diffusion. Around the disk - visible in white - the death halo is apparent, additionally, at the edge of the halo the E. coli bacteria are coloured blueish grey. This shows that as a result of the upregulation of pSoxS in these cells, AmilCP is being produced. This is indicative of a stressful process occurring in these cells. Hereby, we have shown that non-lethal concentrations of nalidixic acid can be detected using disk diffusion with our chromoproteins stress reporter BioBricks.
Our system is designed to selectively respond to antibiotics based on their cellular targets, while not responding to other antibiotics targeting other cellular targets. This allows for early estimations about the mechanism of action of candidate compounds, thereby shortening drug discovery pipelines. Some of the BioBricks we demonstrated this objective with, are pCspA-GFP (BBa_K2610037), pKatG-GFP (BBa_K2610018) and pSoxS-GFP (BBa_K2610031), using flow cytometry. Cells were stressed by a repertoire of known antibiotics at non-lethal concentrations, in order to obtain their unique response profiles, such as can be seen in figure 3A, B and C for pCspA, pSoxS and pKatG respectively. Promoter CspA is suppressed by nearly all antibiotics, but is activated specifically by nalidixic acid. In contrast to pCspA, promoter SoxS is not only activated by nalidixic acid but also by zeocin (both associated with DNA damage). Conversely, promoter KatG is activated only by hydrogen peroxide. Therefore, we have shown that our stress activated promoters react only to specific stressors. This is vitally important for our screening system because it allows for differentiation between modes of actions of candidate compounds.



Combination therapies have been proven to suppress the development of resistance in cancer and HIV treatment[1][2]. Moreover, synergistic combination therapies for fighting infections are already being used in the clinic in the form of amoxicillin/clavulanic acid. During our research we have proven that a non-lethal stressful compound verified by our system could be used to enhance the effect of antibiotics. We screened a number of natural and chemically engineered compounds for their ability to stress bacterial cells, in this case E. coli DH5α. Firstly, compounds that were lethal were eliminated from the screen to ensure results from our system were in response to purely stressful compounds (Figure 4). Consequently, we continued with a select number of compounds due to time restraints. The following compounds were selected through literature research: ginger, ascorbic acid, cinnamon and cannabis[3-8]. We tested the ability of these compounds to activate or suppress various stress-related promoters fused to GFP using flow cytometry.

The clpB promoter (BBa_K2610020), a gene involved in the cellular stress response to aggregated proteins, responded to ascorbic acid, more commonly known as vitamin C (Figure 5). This is an indication that this compound is involved in protein aggregation. Previous research has shown an involvement of ascorbic acid in the inactivation of proteins[9][10]. This suggests our screening system can indicate a possible mechanism of action for stressful compounds.

Ascorbic acid not only emerged as a stressful substance but we also demonstrated that this compound could lower the minimum inhibitory concentration (MIC) of ampicillin (Figure 6A), chloramphenicol (Figure 6B) and nalidixic acid (Figure 6C). This proves our system is able to detect stressful compounds which can be used in antibiotic combination therapies.



References
[1]: Yardley, D. A. (2013). Drug resistance and the role of combination chemotherapy in improving patient outcomes. International journal of breast cancer, 2013.
[2]: Katlama, C., Ingrand, D., Loveday, C., Clumeck, N., Mallolas, J., Staszewski, S., ... & McDade, H. (1996). Safety and efficacy of lamivudine-zidovudine combination therapy in antiretroviral-naive patients: a randomized controlled comparison with zidovudine monotherapy. Jama, 276(2), 118-125.
[3]: Tajkarimi, M., & Ibrahim, S. A. (2011). Antimicrobial activity of ascorbic acid alone or in combination with lactic acid on Escherichia coli O157:H7 in laboratory medium and carrot juice. Food Control, 22(6), 801–804. https://doi.org/10.1016/j.foodcont.2010.11.030
[4]: Mathew, S., Verghese, R., & David, A. (2017). Antimicrobial activity of Vitamin C demonstrated on uropathogenic Escherichia coli and Klebsiella pneumoniae. Journal of Current Research in Scientific Medicine, 3(2), 88. https://doi.org/10.4103/jcrsm.jcrsm_35_17
[5]: Ali, E. M. M., Almagboul, A. Z. I., Khogali, S. M. E., & Gergeir, U. M. A. (2012). Antimicrobial Activity of Cannabis sativa L. Chinese Medicine, 03(01), 61–64. https://doi.org/10.4236/cm.2012.31010
[6]: Chakraborty, S., Afaq, N., Singh, N., & Majumdar, S. (2018). Antimicrobial activity of Cannabis sativa, Thuja orientalis and Psidium guajava leaf extracts against methicillin-resistant Staphylococcus aureus. Journal of Integrative Medicine, 16(5), 350–357. https://doi.org/10.1016/j.joim.2018.07.005
[7]: Patel, R. V., Thaker, V. T., & Patel, V. (2011). Antimicrobial activity of ginger and honey on isolates of extracted carious teeth during orthodontic treatment. Asian Pacific Journal of Tropical Biomedicine, 1(1), 867–871. https://doi.org/10.1016/s2221-1691(11)60124-x
[8]: Gupta, S., & Ravishankar, S. (2005). A Comparison of the Antimicrobial Activity of Garlic, Ginger, Carrot, and Turmeric Pastes Against Escherichia coli O157:H7 in Laboratory Buffer and Ground Beef. Foodborne Pathogens and Disease, 2(4), 330–340. https://doi.org/10.1089/fpd.2005.2.330
[9]: Szarka, A., & Lőrincz, T. (2014). The role of ascorbate in protein folding. Protoplasma, 251(3), 489-497.
[10]: Stadtman, E. R. (1991). Ascorbic acid and oxidative inactivation of proteins. The American journal of clinical nutrition, 54(6), 1125S-1128S.