Difference between revisions of "Team:Queens Canada/Demonstrate"
ElishaKrauss (Talk | contribs) |
JimmyChung (Talk | contribs) |
||
| Line 19: | Line 19: | ||
<div class="column full_size" style="width:70%;margin-left:15%"> | <div class="column full_size" style="width:70%;margin-left:15%"> | ||
<h2>Results</h2> | <h2>Results</h2> | ||
| − | <p>Experiments: | + | <p style="font-size:18px">Experiments: |
This summer QGEM accomplished a lot of work in the laboratory! We have 1) validated the Engineered NanoLuc luciferase as an | This summer QGEM accomplished a lot of work in the laboratory! We have 1) validated the Engineered NanoLuc luciferase as an | ||
choice reporter in our intein-based biosensor construct, 2) We characterized the insolubility of a small-molecule triggered | choice reporter in our intein-based biosensor construct, 2) We characterized the insolubility of a small-molecule triggered | ||
intein splicing system in E. Coli, 3) We validated the nuclear receptor ligand binding as a sensitive domain for analyte | intein splicing system in E. Coli, 3) We validated the nuclear receptor ligand binding as a sensitive domain for analyte | ||
detection through the successful assembly of a reagent-less, and continuous FRET-based biosensor with the ability to discern | detection through the successful assembly of a reagent-less, and continuous FRET-based biosensor with the ability to discern | ||
| − | between agonist and antagonists.<p> | + | between agonist and antagonists.<p style="font-size:18px"> |
<h5>1) Validation of NanoLuc Luciferase for application in our Diagnostic Pacifier</h5> | <h5>1) Validation of NanoLuc Luciferase for application in our Diagnostic Pacifier</h5> | ||
| − | <p>To create our desired portable intein-splicing biosensor, we needed to select a signal that could be measured and correlated | + | <p style="font-size:18px">To create our desired portable intein-splicing biosensor, we needed to select a signal that could be measured and correlated |
to the amount of analyte present. Additionally, we had the challenge of selecting a reporter that could be produced and quantified | to the amount of analyte present. Additionally, we had the challenge of selecting a reporter that could be produced and quantified | ||
very portably. Our first idea was to use a fluorescent reporter, however with fluorescence, a fluorophore must first be excited by | very portably. Our first idea was to use a fluorescent reporter, however with fluorescence, a fluorophore must first be excited by | ||
| Line 57: | Line 57: | ||
<img style="margin-left:15%" src="https://static.igem.org/mediawiki/2018/f/f9/T--Queens_Canada--NanoTimelapse.jpeg" width="70%"/> | <img style="margin-left:15%" src="https://static.igem.org/mediawiki/2018/f/f9/T--Queens_Canada--NanoTimelapse.jpeg" width="70%"/> | ||
| − | <p>We next sough to characterize the sensitivity of our portable luminometer to the luminescence produced by NanoLuc. The portable | + | <p style="font-size:18px">We next sough to characterize the sensitivity of our portable luminometer to the luminescence produced by NanoLuc. The portable |
luminometer measures differently than our labs luminometer, as the Lumistar Glaxay luminometer in our lab takes an instantaneous | luminometer measures differently than our labs luminometer, as the Lumistar Glaxay luminometer in our lab takes an instantaneous | ||
measurement of light, in relative light units. Whereas, our built luminometer measures photons/second. Therefore, the two devices | measurement of light, in relative light units. Whereas, our built luminometer measures photons/second. Therefore, the two devices | ||
| Line 73: | Line 73: | ||
| − | <p> The goal of this year’s project was to create a biosensor that would luminesce in the presence of cortisol, giving our team the ability to quantify the hormone in saliva. The idea was ignited from an small-molecule triggered intein previously developed by Buskirk et al (2004). In this article, the authors inserted the ligand binding domain of the Estrogen Receptor into the RecA intein, disrupting its ability to perform auto-splicing, as found in nature. Splicing was only restored after the addition of the estrogen antagonist 4-hydroxytamoxifen (4-HT), and rounds of mutagenesis. The construct was then inserted into variety of split extein contexts, such as kanamycin resistance, and GFP, and found to splice together the split proteins halves and produce the mature protein, in a dose-responsive manner to 4-HT addition. | + | <p style="font-size:18px"> The goal of this year’s project was to create a biosensor that would luminesce in the presence of cortisol, giving our team the ability to quantify the hormone in saliva. The idea was ignited from an small-molecule triggered intein previously developed by Buskirk et al (2004). In this article, the authors inserted the ligand binding domain of the Estrogen Receptor into the RecA intein, disrupting its ability to perform auto-splicing, as found in nature. Splicing was only restored after the addition of the estrogen antagonist 4-hydroxytamoxifen (4-HT), and rounds of mutagenesis. The construct was then inserted into variety of split extein contexts, such as kanamycin resistance, and GFP, and found to splice together the split proteins halves and produce the mature protein, in a dose-responsive manner to 4-HT addition. |
We sought to expand on the detection capabilities of this system, by creating a modular system, where both the ligand binding domain, and the extein could be replaced. The ultimate goal of the project was to create a biological sensor for the detection of cortisol. We would achieve this by swapping the estrogen receptor ligand binding domain for the glucocorticoid receptor, and replace the kanamycin resistance extein with the bright luciferase, NanoLuc. | We sought to expand on the detection capabilities of this system, by creating a modular system, where both the ligand binding domain, and the extein could be replaced. The ultimate goal of the project was to create a biological sensor for the detection of cortisol. We would achieve this by swapping the estrogen receptor ligand binding domain for the glucocorticoid receptor, and replace the kanamycin resistance extein with the bright luciferase, NanoLuc. | ||
<br><br> | <br><br> | ||
| Line 94: | Line 94: | ||
</p> | </p> | ||
| − | <p> | + | <p style="font-size:18px"> |
We then transformed our construct into BL-21 DE3, a strain of E. coli that is important for protein expression, and induced expression with IPTG. To test intein splicing, we made agar plates containing either 1) Ampicillin, as a positive control, 2) Kanamycin, as a negative control 3) Kanamycin and 4-HT. We plated the BL21 DE3 and found that no bacterial colonies grew on the Kanamycin alone, or Kanamycin and 4-HT containing plates, but there was growth on the Ampicillin containing plates. If the iintein was in fact splicing caused by 4-HT, we would expect to see growth on the Kanamycin and 4-HT containing plate. Therefore, this demonstrated that the intein construct was not splicing and Kanamycin Resistance was not being produced. | We then transformed our construct into BL-21 DE3, a strain of E. coli that is important for protein expression, and induced expression with IPTG. To test intein splicing, we made agar plates containing either 1) Ampicillin, as a positive control, 2) Kanamycin, as a negative control 3) Kanamycin and 4-HT. We plated the BL21 DE3 and found that no bacterial colonies grew on the Kanamycin alone, or Kanamycin and 4-HT containing plates, but there was growth on the Ampicillin containing plates. If the iintein was in fact splicing caused by 4-HT, we would expect to see growth on the Kanamycin and 4-HT containing plate. Therefore, this demonstrated that the intein construct was not splicing and Kanamycin Resistance was not being produced. | ||
<br><br> | <br><br> | ||
| Line 105: | Line 105: | ||
<br><br> | <br><br> | ||
</p> | </p> | ||
| − | <p>To discern why the intein was not splicing we ran a SDS-page on induced BL21-DE3 expression. We grew 2 liquid cultures of BL21 DE3 containing the intein, induced expression of T7 RNA polymerase with IPTG, then incubated both for 4 hours at 37C. We then added 10μm 4-HT to one of the liquid cultures and incubated at 37C for an additional 1 hour to allow splicing of the intein to occur.<br> | + | <p style="font-size:18px">To discern why the intein was not splicing we ran a SDS-page on induced BL21-DE3 expression. We grew 2 liquid cultures of BL21 DE3 containing the intein, induced expression of T7 RNA polymerase with IPTG, then incubated both for 4 hours at 37C. We then added 10μm 4-HT to one of the liquid cultures and incubated at 37C for an additional 1 hour to allow splicing of the intein to occur.<br> |
| − | </p><p> | + | </p><p style="font-size:18px"> |
The team lysed the cells by addition of lysozyme and protease inhibitor containing lysis buffer, followed by sonication for 10 seconds. 10 seconds would provide sufficient enough time for splicing without destroying the cells, or having the liquid bacteria overheat. We then collected 1ml of the mixed fraction and stored at -20C. The remainder of the fraction were centrifuged at 8k RPM for 5 minutes and collected 1ml of the soluble fraction. The remainder of the soluble fraction was discarded, and the insoluble fraction was resuspended, and 1 ml was collected as the insoluble fraction. | The team lysed the cells by addition of lysozyme and protease inhibitor containing lysis buffer, followed by sonication for 10 seconds. 10 seconds would provide sufficient enough time for splicing without destroying the cells, or having the liquid bacteria overheat. We then collected 1ml of the mixed fraction and stored at -20C. The remainder of the fraction were centrifuged at 8k RPM for 5 minutes and collected 1ml of the soluble fraction. The remainder of the soluble fraction was discarded, and the insoluble fraction was resuspended, and 1 ml was collected as the insoluble fraction. | ||
<br> | <br> | ||
| Line 116: | Line 116: | ||
</figcaption> | </figcaption> | ||
</figure> | </figure> | ||
| − | <br></p><p> | + | <br></p><p style="font-size:18px"> |
The SDS page informed us that the majority of our protein product was found in the pellet and not the supernatant. This indicated that our construct was not soluble, and would not be able to splice. | The SDS page informed us that the majority of our protein product was found in the pellet and not the supernatant. This indicated that our construct was not soluble, and would not be able to splice. | ||
We subsequently found one paper that had previously utilized dose-dependent intein splicing in bacteria, however this paper further tagged intein constructs with a maltose-binding protein.[2] Therefore, this further demonstrates the 4-HT dependent intein to be insoluble in Bacteria, unless its solubility is drastically increased through the use of solubility tags such as Maltose-binding protein. | We subsequently found one paper that had previously utilized dose-dependent intein splicing in bacteria, however this paper further tagged intein constructs with a maltose-binding protein.[2] Therefore, this further demonstrates the 4-HT dependent intein to be insoluble in Bacteria, unless its solubility is drastically increased through the use of solubility tags such as Maltose-binding protein. | ||
Revision as of 23:56, 15 October 2018
Results
Experiments: This summer QGEM accomplished a lot of work in the laboratory! We have 1) validated the Engineered NanoLuc luciferase as an choice reporter in our intein-based biosensor construct, 2) We characterized the insolubility of a small-molecule triggered intein splicing system in E. Coli, 3) We validated the nuclear receptor ligand binding as a sensitive domain for analyte detection through the successful assembly of a reagent-less, and continuous FRET-based biosensor with the ability to discern between agonist and antagonists.
1) Validation of NanoLuc Luciferase for application in our Diagnostic Pacifier
To create our desired portable intein-splicing biosensor, we needed to select a signal that could be measured and correlated
to the amount of analyte present. Additionally, we had the challenge of selecting a reporter that could be produced and quantified
very portably. Our first idea was to use a fluorescent reporter, however with fluorescence, a fluorophore must first be excited by
a light, and then a light is given off that needs to filter and its intensity measured. Fluorimeter are bulky devices, and after
further investigation, it become apparent that it would be possible the machinery down to fit with a pacifier. However, during our
investigations we learned about the elements needed to build a luminometer and found that it would be entirely feasible for us to
produce a simplified photon-counter device that could fit into a pacifier! (See Hardware section).
To best improve our chances of being able to produce a signal which our portable luminometer could detect we investigated various
the relative strength of luciferases. We ultimately decided on testing the novel and very bright NanoLuc Luciferase as the quantitative
reporter for our construct. NanoLuc® developed by Promega is a luciferase derived via directed evolution from the luminous shrimp,
Oplophorus gracilirostris [4]. The enzyme was obtained from deep-sea shrimp and optimized following the discovery of a novel substrate,
furimazine, which allows for the production of visible light with less background activity than other luciferases [4,5]. NanoLuc® is a
relatively small, a19.1 kDa monomeric protein that is both soluble and ATP-independent [4]. Compared to firefly (Lampyridae) and sea
pansy (Renilla) luciferases, this novel protein offers many advantages reflected by its increased stability, smaller size, and >150-fold
increase in luminescence [5]. The unique characteristics of this enzyme construct combined with its high luminescence activity allow for
the production of a very sensitive diagnostic assay.
By generous donation from Promega, we received the promoter-less vector pNL1.1. We then cloned an Anderson Constitutive promoter
(Part:BBa_J23100) into the MCS of pNL1.1. To test our construct, we grew an overnight liquid culture of E. Coli expressing the plasmid
construct. We pelleted 2-3ml of liquid culture, resuspended in a lytic buffer with lysozyme and protease inhibitor for 30 mins, before
sonicating for 10 seconds the placing on ice, twice. We then pelleted the insoluble fraction, collected the supernatant and performed a
Luciferase Assay, using Nano-Glo® Luciferase Assay System. Relative light units were measured in a Lumistar Galaxy plate reader. We
then took an initial measurement of Relative Light Unit and found a strong result of 38598.2±3385.40 RLU (n=4), compared to 103.5±33.6
RLU (n=5), with our negative control at peak luminescence. Since the ultimate goal of using NanoLuc will be to produce a signal for
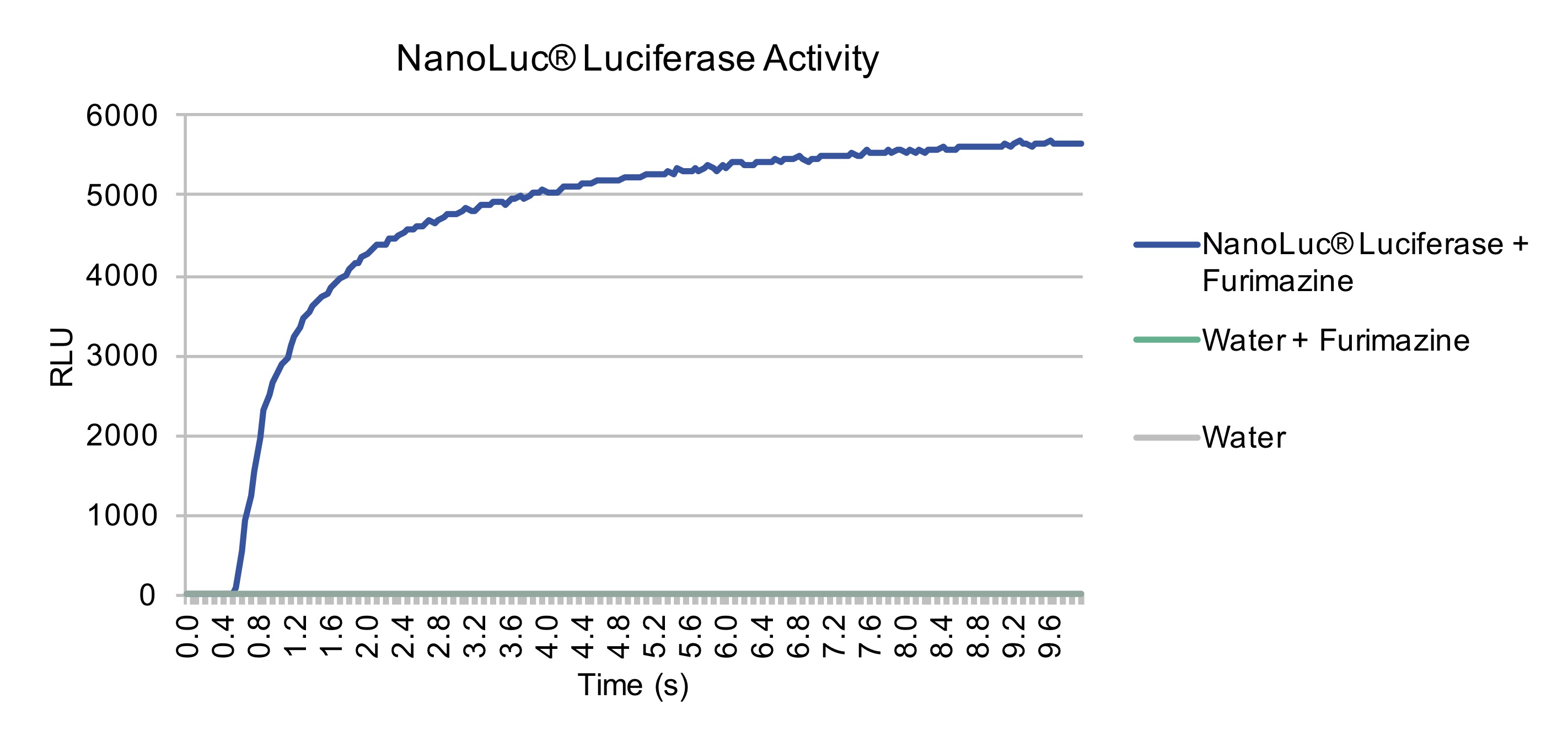
diagnostic purposes, we sought to further characterize the time-course of NanoLuc Luciferase after the addition of substrate. Through
taking measurements every 0.05 seconds, we were able to produce a curve demonstrating the time till plateau of NanoLuc after addition
of substrate. Within 10 seconds, NanoLuc had reached its plateau, therefore validating NanoLuc would be an excellent reporter protein
for application in our intein-based diagnostic biosensor.
We next sough to characterize the sensitivity of our portable luminometer to the luminescence produced by NanoLuc. The portable luminometer measures differently than our labs luminometer, as the Lumistar Glaxay luminometer in our lab takes an instantaneous measurement of light, in relative light units. Whereas, our built luminometer measures photons/second. Therefore, the two devices cannot be directly compared. For calibration of our luminometer we first confirmed the sample to be luminescing by a measurement with the Lumistar Galaxy Luminometer, we then measured samples diluted 1, 2, and 4-fold and found our luminometer was capable of measuring a linear relationship between the concentrations of our sample. Thus confirming our luciferase construct was functional and rised to plateau quickly, our built portable luminometer was accurate across a range of concentrations, and that Nanoluc Luciferase could serve as a viable signal for our diagnostic assays.

2) Characterization of insolubility of a small-molecule triggered intein splicing system in E. Coli
The goal of this year’s project was to create a biosensor that would luminesce in the presence of cortisol, giving our team the ability to quantify the hormone in saliva. The idea was ignited from an small-molecule triggered intein previously developed by Buskirk et al (2004). In this article, the authors inserted the ligand binding domain of the Estrogen Receptor into the RecA intein, disrupting its ability to perform auto-splicing, as found in nature. Splicing was only restored after the addition of the estrogen antagonist 4-hydroxytamoxifen (4-HT), and rounds of mutagenesis. The construct was then inserted into variety of split extein contexts, such as kanamycin resistance, and GFP, and found to splice together the split proteins halves and produce the mature protein, in a dose-responsive manner to 4-HT addition.
We sought to expand on the detection capabilities of this system, by creating a modular system, where both the ligand binding domain, and the extein could be replaced. The ultimate goal of the project was to create a biological sensor for the detection of cortisol. We would achieve this by swapping the estrogen receptor ligand binding domain for the glucocorticoid receptor, and replace the kanamycin resistance extein with the bright luciferase, NanoLuc.
Nuclear receptors such as estrogen and glucocorticoid share large homology in their DNA sequence, and 3D strucutre. The generic LBD of nuclear receptors is comprised of 12 a-helices and 4 small β-sheets that fold into a three-layer helical domain. Upon binding with an antagonist or agonist, the receptors undergo a conformational change. The conformational change is highlighted in the movement of helix 12. Our team modeld a variety of linkers for the glucocorticoid, which would connect the halves of the RecA intein upon cortisol binding We wanted linkers long enough to be able to wrap around the molecule upon conformational change and binding with the correct ligand, however the linkers needed to be rigid and short enough, not to induce auto-splicing without the need of ligand binding. The following linkers were designed based on the optimal angstrom distances and flexible design. Our dry lab team calculated that for a rigid linker, the shortest possible distance needed when the conformational change was initiated was 8 angstrom, and 20 angstrom for a more flexible linkers. We tried out three different linkers for our glucocorticoid construct, as we were unsure how rigid or flexible the linker needed to be to ensure splicing. Once the linkers were chosen, we performed molecular modelling to ensure that the two inteins halves would come into close enough contact to initiate splicing, and thus produce luminescence. The molecular dynamic simulations verified that the linkers: N-terminal: GPGSGS, C-terminal: GASGSG, were in fact long enough for the intein to come in contact, and they even formed a favourable interaction in just 10 nanosecond of simulation time. (see Molecular Dynamic Simulations her). Armed with knowledge gained in our dry lab, we set out to recreate the results by Buskirk et al. (2004), and develop new constructs based on the glucocorticoid receptor.
For proof of concept, we conducted experiments on these constructs in Eschiceria coli. Although the original paper utilized yeast, we set out to test the ability of the 4-HT dependent intein to splice in bacteria, in efforts to improve its characterization, and due to the relative ease of working with novel genetic constructs in yeast. We firstly constructed the Kanamycin-4HT-Intein, and Kanamycin-GR-Intein in the expression vector pET16b via gibson assembely with overlapping sequence, and then transformed DH5-alpha. We picked colonies, grew liquid cultures, mini-prepped, and performed diagnostic digests to confirm our insert was present.


We then transformed our construct into BL-21 DE3, a strain of E. coli that is important for protein expression, and induced expression with IPTG. To test intein splicing, we made agar plates containing either 1) Ampicillin, as a positive control, 2) Kanamycin, as a negative control 3) Kanamycin and 4-HT. We plated the BL21 DE3 and found that no bacterial colonies grew on the Kanamycin alone, or Kanamycin and 4-HT containing plates, but there was growth on the Ampicillin containing plates. If the iintein was in fact splicing caused by 4-HT, we would expect to see growth on the Kanamycin and 4-HT containing plate. Therefore, this demonstrated that the intein construct was not splicing and Kanamycin Resistance was not being produced.

To discern why the intein was not splicing we ran a SDS-page on induced BL21-DE3 expression. We grew 2 liquid cultures of BL21 DE3 containing the intein, induced expression of T7 RNA polymerase with IPTG, then incubated both for 4 hours at 37C. We then added 10μm 4-HT to one of the liquid cultures and incubated at 37C for an additional 1 hour to allow splicing of the intein to occur.
The team lysed the cells by addition of lysozyme and protease inhibitor containing lysis buffer, followed by sonication for 10 seconds. 10 seconds would provide sufficient enough time for splicing without destroying the cells, or having the liquid bacteria overheat. We then collected 1ml of the mixed fraction and stored at -20C. The remainder of the fraction were centrifuged at 8k RPM for 5 minutes and collected 1ml of the soluble fraction. The remainder of the soluble fraction was discarded, and the insoluble fraction was resuspended, and 1 ml was collected as the insoluble fraction.
We then ran a SDS-page to determine protein expression, and if splicing had occurred.

The SDS page informed us that the majority of our protein product was found in the pellet and not the supernatant. This indicated that our construct was not soluble, and would not be able to splice.
We subsequently found one paper that had previously utilized dose-dependent intein splicing in bacteria, however this paper further tagged intein constructs with a maltose-binding protein.[2] Therefore, this further demonstrates the 4-HT dependent intein to be insoluble in Bacteria, unless its solubility is drastically increased through the use of solubility tags such as Maltose-binding protein.
Our next step was to try and increase our protein’s solubility. Through a collaboration with the Stonybrook iGEM team, we decided to try directed evolution through error-prone PCR. We created primers for the insert and the backbone, and added MnCl¬2 to our PCR reaction to induce random mutagenesis. We then tried Gibson assembly and transformation of the mutagenized product to see if any insert or backbone had worked. If colonies grew on kanamycin and cortisol, we know that splicing worked. The colonies would then be sampled and plated on kanamycin, if they grew, then the intein was not selective for just ligand binding. Unfortunately, we saw no evidence of the error prone PCR being successful after numerous attempts.
In the future, our constructs solubility could be increased through error-prone PCR, with a careful selection process. As well, we discussed the possibility of adding a maltose binding domain to our protein and increasing its solubility. E. coli maltose binding protein (MBP), can be used to promote solubility of the protein that it is fused with.
Since the intein construct previously worked in yeast, we also believe our construct could be successfully constructed and spliced upon ligand binding in this organism.
3) Validation of the nuclear receptor ligand binding as a ligand binding domain producing pronounced conformational changes through the successful assembly of a reagent-less, and continuous FRET-based biosensor with the ability to discern between agonist and antagonists.
References
- A. R. Buskirk, Y.-C. Ong, Z. J. Gartner, and D. R. Liu," “Directed evolution of ligand dependence: small-molecule-activated protein splicing., Proc. Natl. Acad. Sci. U. S. A., vol. 101, no. 29, pp. 10505–10, Jul. 2004.
- C. G. England, E. B. Ehlerding, and W. Cai, “NanoLuc: A Small Luciferase Is Brightening Up the Field of Bioluminescence., Bioconjug. Chem., vol. 27, no. 5, pp. 1175–1187, 2016."
- . Boute, P. Lowe, S. Berger, M. Malissard, A. Robert, and M. Tesar, “NanoLuc Luciferase - A Multifunctional Tool for High Throughput Antibody Screening., Front. Pharmacol., vol. 7, p. 27, 2016.
- L. G. G. C. Verhoef, M. Mattioli, F. Ricci, Y.-C. Li, and M. Wade, “Multiplex detection of protein–protein interactions using a next generation luciferase reporter,” Biochim. Biophys. Acta - Mol. Cell Res., vol. 1863, no. 2, pp. 284–292, Feb. 2016.