Difference between revisions of "Team:Bielefeld-CeBiTec/Software"
| Line 98: | Line 98: | ||
<div class="title">siRCon - A siRNA Constructor</div> | <div class="title">siRCon - A siRNA Constructor</div> | ||
| + | <h2>Short Summary</h2> | ||
<article> | <article> | ||
| − | In our project we introduce RNA interference (RNAi) and translation repression with small interfering ( | + | In our project, we introduce RNA interference (RNAi) and translation repression with small interfering RNAs (siRNAs) as an alternative to CRISPR/Cas. To use siRNA as silencing agents for the gene-of -interest, we propose a two-step design process. At first, potential siRNAs for prokaryotic organisms must be designed. In the second step, the silencing effect of these siRNAs can be validated by our siRNA vector system <a href="https://2018.igem.org/Team:Bielefeld-CeBiTec/siRNA">Tace.</a> To facilitate the initial siRNA design step, we developed a siRNA construction tool which identifies possible siRNAs for a given gene sequence, calculates their probability to silence the target gene, and delivers candidates ranked based on the calculated score. It consists of three modules: "siRNAs for RNAi", "siRNA", and "check siRNA". The siRNAs predicted by our software are perfectly compatible with our siRNA vector system. To the best of our knowledge, this is the first tool dedicated to predicting customized siRNA for application in prokaryotes. This Python tool comes in two versions: a command line application and an easy-to-use graphical interface. |
</article> | </article> | ||
<a name="intro" id="intro" class="shifted-anchor"></a> | <a name="intro" id="intro" class="shifted-anchor"></a> | ||
| − | <h2> | + | <h2>siRNAs short introduction</h2> |
<article> | <article> | ||
| − | siRNAs are small single | + | siRNAs are small, non-coding single-stranded RNAs with an average length of 21-25 nucleotidesnucleotides which bind a specific complementary coding mRNA and silence its function. In eukaryotic RNAi, siRNAs are loaded to Argonaute proteins which carry out the repression, either by blocking mRNA translation or by degrading the mRNA (Siomi and Siomi, 2009). More detailed information on both possible siRNAs mechanisms is found <a href="https://2018.igem.org/Team:Bielefeld-CeBiTec/siRNA">here.</a> |
</article> | </article> | ||
<a name="overh" id="overh" class="shifted-anchor"></a> | <a name="overh" id="overh" class="shifted-anchor"></a> | ||
| − | <h2>siRNA | + | <h2>siRNA design</h2> |
<article> | <article> | ||
| − | In order to achieve effective gene silencing or knockdown, the 19 nt binding sequence must be | + | In order to achieve effective gene silencing or knockdown, the 19 nt binding sequence must be flanked by special, non-binding 5' and 3' extensions (Figure 1). To trigger mRNA degradation by RNase E, the 5’-terminal triphosphate of the siRNA needs to be converted to a monophosphate by RNA pyrophosphohydrolase (RppH). For the siRNA to be recognized by RppH, the 5’ end of the siRNA has to start with the tetranucleotide AGNN which is not allowed to match the targeted mRNA (Foley et al., 2015). At the 3’ end of the siRNA, the small MicC scaffold is added which facilitates the hybridization of siRNA and target mRNA and protects the siRNA from degradation (Na et al., 2013). |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
</article> | </article> | ||
| Line 123: | Line 120: | ||
<img class="figure hundred" src="https://static.igem.org/mediawiki/2018/9/9a/T--Bielefeld-CeBiTec--RNAi_scaffolds_new.png"> | <img class="figure hundred" src="https://static.igem.org/mediawiki/2018/9/9a/T--Bielefeld-CeBiTec--RNAi_scaffolds_new.png"> | ||
<figcaption> | <figcaption> | ||
| − | <b>Figure 1:</b> | + | <b>Figure 1:</b> Effects of siRNA design on RNAi effectiveness and siRNA stability. <b>A</b> If the siRNA does not carry suitable 5' or 3' extensions, it is quickly degraded. <b>B</b> siRNAs extended by the tetranucleotide AGNN are recognized and processed by the pyrophosphohydrolase RppH. This enzyme converts the 5' triphosphate to a monophosphate which greatly reduces siRNA degradation. This allows the siRNA to hybridize to its target mRNA which in turn is degraded by RNAse E, thus leading to effective mRNA silencing. <b>C</b> Extending siRNAs with a 3' MicC scaffold in addition to the 5' tetranucleotide AGNN further enhances mRNA silencing. MicC facilitates the hybridization of siRNA and target mRNA and protects the siRNA from degradation. |
</figcaption> | </figcaption> | ||
</figure> | </figure> | ||
| Line 129: | Line 126: | ||
<article> | <article> | ||
| − | + | In addition to degredation-based RNAi, siRNA can also be used to block mRNAs without degradation. This is achieved by adding the outer membrane protein A (OmpA) scaffold to the 5' end of the siRNA (Figure 2), enhancing its stability. In addition, the hybridization of the siRNA and the target mRNA can be facilitated by addition of MicC to the 3' terminus. | |
</br> | </br> | ||
| − | + | Both sequence extensions are also part of our vector system, enabling efficient design and construction of effective siRNAs. If our vector system is selected when using our tool, the fitting overlaps to our vectors are added automatically. More theoretical information about the overhangs and scaffolds can be found <a href="">here</a>. | |
</article> | </article> | ||
| Line 137: | Line 134: | ||
<img class="figure sixty" src="https://static.igem.org/mediawiki/2018/f/f0/T--Bielefeld-CeBiTec--siRNA_scaffolds_new_vk.png"> | <img class="figure sixty" src="https://static.igem.org/mediawiki/2018/f/f0/T--Bielefeld-CeBiTec--siRNA_scaffolds_new_vk.png"> | ||
<figcaption> | <figcaption> | ||
| − | <b>Figure 2:</b> | + | <b>Figure 2:</b> siRNA design for silencing translation. <b>A</b> If the siRNA does not carry suitable 5' or 3' extensions, it is quickly degraded. <b>B</b> siRNAs supplemented with the outer membrane protein A (OmpA) scaffold are more stable and effectively silence the translation of target mRNAs. <b>C</b> If the siRNA is supplemented with the OmpA as well as the MicC scaffold the repression is enhanced further. </figcaption> |
</figure> | </figure> | ||
| Line 145: | Line 142: | ||
<article> | <article> | ||
| − | In 2012 the <a href="https://2012.igem.org/Team:SYSU-Software/Models#pp2">iGEM team SYSU-Software</a> integrated an siRNA cDNA designer as a small part of their project. siRNAs designed with this tool were applicable in eukaryotic organisms. They included two different design methods: Tom Tuschl’s method and Rational siRNA design. | + | In 2012, the <a href="https://2012.igem.org/Team:SYSU-Software/Models#pp2">iGEM team SYSU-Software</a> integrated an siRNA cDNA designer as a small part of their project. siRNAs designed with this tool were applicable in eukaryotic organisms. They included two different design methods: Tom Tuschl’s method and Rational siRNA design. |
</article> | </article> | ||
| Line 156: | Line 153: | ||
<article> | <article> | ||
| − | Tom Tuschl’s method focuses mainly on the existence of 5’ and 3’ ‘TT’ overhangs (Figure 3)(Elbashir et al., 2001). These are not compatible with overhangs and scaffold sequences | + | Tom Tuschl’s method focuses mainly on the existence of 5’ and 3’ ‘TT’ overhangs (Figure 3) (Elbashir <i>et al.</i>, 2001). These are not compatible with overhangs and scaffold sequences required by the prokaryotic mechanisms. Therefore, we decided to use the rules published by Ui-Tei as an alternative design method (Naito and Ui-Tei, 2012). Furthermore, we adapted the Rational siRNA design as it was more suitable for our application (Reynolds <i>et al.</i>, 2004). Both design rules apply only to the 19 nt long target binding sequence. |
</article> | </article> | ||
| Line 163: | Line 160: | ||
<article> | <article> | ||
| − | By a systematic analysis of 180 eukaryotic siRNAs Reynolds et al. identified eight criteria that are important for their functionality (Reynolds et al., 2004). Each criterion gets a score that is either positive or negative, corresponding to its effect on the siRNA. All siRNA candidates | + | By a systematic analysis of 180 eukaryotic siRNAs, Reynolds <i>et al.</i> identified eight criteria that are important for their functionality (Reynolds et al., 2004). Each criterion gets a score that is either positive or negative, corresponding to its effect on the siRNA. All siRNA candidates with a score above six are potential highly functional siRNAs. |
</article> | </article> | ||
| Line 177: | Line 174: | ||
</tr> | </tr> | ||
<tr> | <tr> | ||
| − | <td>At least 3 'A | + | <td>At least 3 'W' ('A' or 'T') at positions 15-19</td> |
<td>+1 (for each 'A' or 'T')</td> | <td>+1 (for each 'A' or 'T')</td> | ||
</tr> | </tr> | ||
| Line 197: | Line 194: | ||
</tr> | </tr> | ||
<tr> | <tr> | ||
| − | <td>An 'A' or 'T' at | + | <td>An 'A' or 'T' at position 19</td> |
<td>-1</td> | <td>-1</td> | ||
</tr> | </tr> | ||
| Line 226: | Line 223: | ||
<li>A ‘G’ or ‘C’ at position 1</li> | <li>A ‘G’ or ‘C’ at position 1</li> | ||
<li>At least five ‘T’ or ‘A’ residues from positions 13 to 19</li> | <li>At least five ‘T’ or ‘A’ residues from positions 13 to 19</li> | ||
| − | <li>No ‘GC’ stretch more than | + | <li>No ‘GC’ stretch more than 9 nt long</li> |
</ol> | </ol> | ||
| Line 233: | Line 230: | ||
<article> | <article> | ||
| − | + | Our software siRCon should report not only the sequences of potential effective siRNAs, but also rank them based on the probability with which they are effective. This is calculated with the help of Bayes’ theorem by calculating probabilities of dependent events. The following calculations and formulas are based on Takasaki (2009). | |
</br> | </br> | ||
| − | + | TThe initial hypothesis is that the given siRNA effectively silences an mRNA. To perform the calculations, a prior probability is necessary. The prior probability for effective gene silencing of mammalian genes can be obtained from former siRNA experiments and is approximately 0.1 (Takasaki, 2009). Since we have no data on prokaryotic siRNAs, we use the same prior probability for our predictions. </br> | |
| − | The gene silencing probability | + | The gene silencing probability \(P(eff|X)\) is described as: |
| − | $$ P(eff|X) = \frac{P^{eff} P(X|eff)}{P(X)} \qquad (1)$$ | + | $$ P(eff|X) = \frac{P^{eff} P(X|eff)}{P^{eff} P(X|eff) + P^{inf} P(X|inf)} \qquad (1)$$ |
| − | + | The 19 nt siRNA binding sequence is represented by X, where \(x_i^n\) corresponds to the bases adenine, guanine, cytosine or thymine (indexes 1≤n≤4) at sequence position i. The probabilities P(X|eff) and P(X|inf) are calculated based on prior knowledge about siRNA sequences that were shown to be effective respectively ineffective in silencing their target mRNAs. Based on the analysis of 833 effective and 847 ineffective siRNAs, Takasaki et al. determined the likelyhood with which base n occures at position i in an effective/ineffective siRNA sequences, represented by the coefficients \(q_{x_i^n}^{eff}\) and \(q_{x_i^n}^{inf}\) respectively. These coeffecients are often referred to as frequency ratios of n at position i. | |
| − | + | ||
| − | + | ||
| − | + | ||
</article> | </article> | ||
<article> | <article> | ||
| − | \( | + | \(P(X|eff)\) and \(P(X|inf)\) are computed as the product of the frequency ratios for each base n at position i in the siRNA binding sequence: |
| − | + | ||
| − | + | ||
| − | + | $$ P(X|inf) = \prod_{i=1}^{19} q_{x_i^n}^{eff} \qquad (2)$$ | |
| − | + | $$ P(X|inf) = \prod_{i=1}^{19} q_{x_i^n}^{inf} \qquad (3)$$ | |
| − | + | ||
| − | $$ P(X|inf) = \prod_{i=1}^{19} q_{x_i^n}^{inf} \qquad ( | + | |
</article> | </article> | ||
| − | |||
<article> | <article> | ||
| − | + | Both probabilities are weighted with their prior probabilities, \(P^{eff}\) and \(P^{inf} = 1-P^{eff}\), where \(P^{eff}\) is set to 0.1 as mentioned previously. With all defined formulas (1),(2) and (3), the gene silencing probability \(P(eff|X)\) is calculated as follows: | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | With all defined formulas | + | |
$$P(eff|X) = \frac{P^{eff} P(X|eff)}{P^{eff} P(X|eff)+P^{inf} P(X|inf)} \\\\= \frac{P^{eff} \prod_{i=1}^{19} q_{x_i^n}^{eff}}{P^{eff} \prod_{i=1}^{19} q_{x_i^n}^{eff}+P^{inf} \prod_{i=1}^{19} q_{x_i^n}^{inf}} $$ | $$P(eff|X) = \frac{P^{eff} P(X|eff)}{P^{eff} P(X|eff)+P^{inf} P(X|inf)} \\\\= \frac{P^{eff} \prod_{i=1}^{19} q_{x_i^n}^{eff}}{P^{eff} \prod_{i=1}^{19} q_{x_i^n}^{eff}+P^{inf} \prod_{i=1}^{19} q_{x_i^n}^{inf}} $$ | ||
| − | |||
</article> | </article> | ||
| Line 277: | Line 261: | ||
<article> | <article> | ||
| − | + | For each nucleotide, the probability of occurrence was determined for each position of the siRNA. Different models were taken into account in the calculation. First of all, the occurrence of the different nucleotides at positions 1 to 19 can be considered as independent of each other. The probabilities for each position are then calculated independently. However, the occurrences of the nucleotides can also be considered as dependent of each other. This means that the occurrence of a nucleotide depends on the nucleotide at the position before. For the calculation of dependent probabilities, the Simple Markow Model was used. It has been found that the resulting silence probability is most accurate when the frequency ratios of the effective siRNAs are calculated dependent and the frequency ratios of the ineffective siRNAs are calculated independent. All frequency ratios can be looked up | |
| + | <a href="https://static.igem.org/mediawiki/2018/5/51/T--Bielefeld-CeBiTec--frequency_ratios_vk.pdf" style="padding-right:0; margin-right:0;">here</a>. | ||
</br> | </br> | ||
| − | In combination with the frequency ratios it is now possible to calculate the silencing probability for the 19 bp long binding site of siRNAs. | + | In combination with the frequency ratios it is now possible to calculate the silencing probability for the 19 bp long binding site of siRNAs. |
</article> | </article> | ||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
Revision as of 01:44, 18 October 2018

siRCon - A siRNA Constructor
Short Summary
siRNAs short introduction
siRNA design


Choosing appropriate design methods

Rational siRNA design
| Rule | Score |
|---|---|
| 30%-52% G/C content | +1 |
| At least 3 'W' ('A' or 'T') at positions 15-19 | +1 (for each 'A' or 'T') |
| Absence of internal repeats (\(T_m \lt 20\)) | +1 |
| An 'A' at position 3 | +1 |
| An 'A' at position 19 | +1 |
| A 'T' at position 19 | +1 |
| An 'A' or 'T' at position 19 | -1 |
| An 'A', 'C' or 'T' at position 13 | -1 |
Ui-Tei rule
- An ‘A’ or ‘T’ at position 19
- A ‘G’ or ‘C’ at position 1
- At least five ‘T’ or ‘A’ residues from positions 13 to 19
- No ‘GC’ stretch more than 9 nt long
Calculating silencing probability
siRNA construction for RNAi and repression of translation
Check siRNA
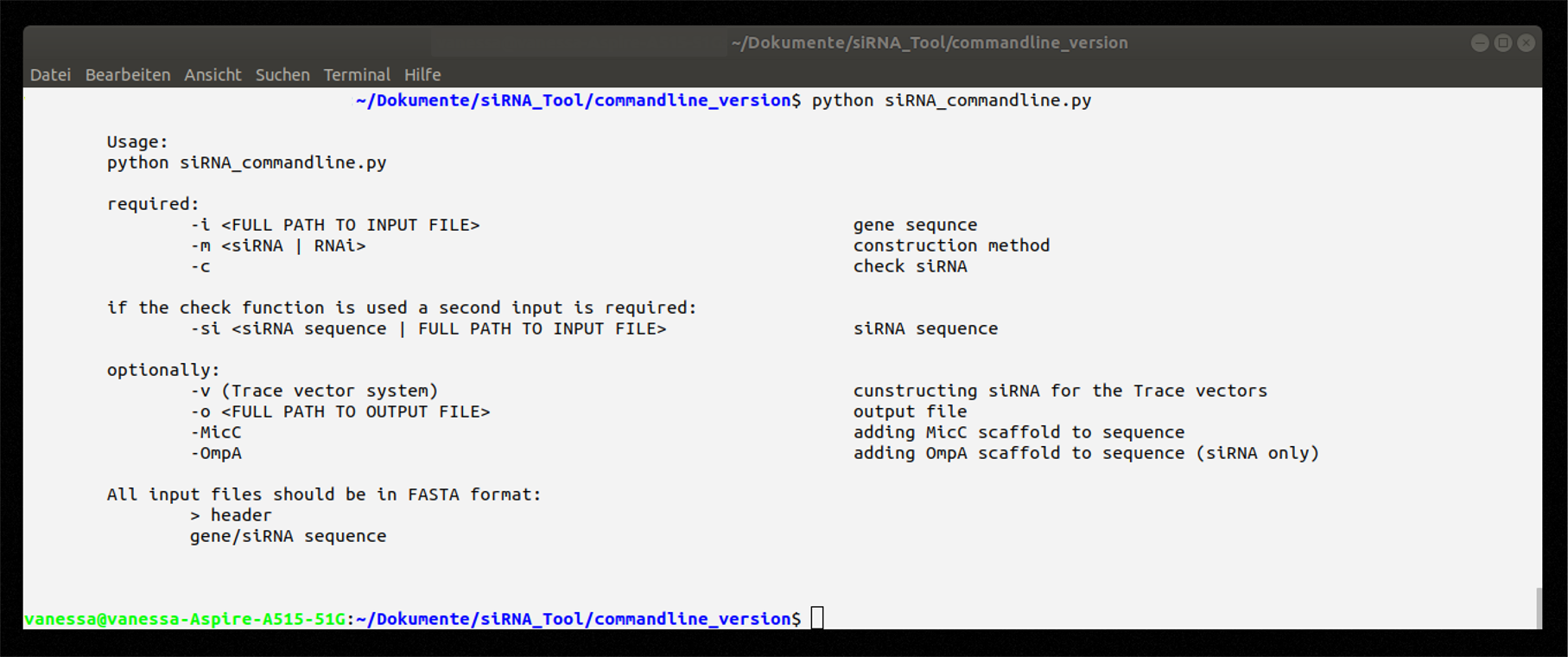
Command line application
The command line application can be obtained directly here or downloaded from our GitHub repository. For the execution of this command line application Python 2.7 needs to be installed.

Graphical Interface usage
As the command line application, the graphical interface version can either be downloaded directly here, or via our GitHub repository.
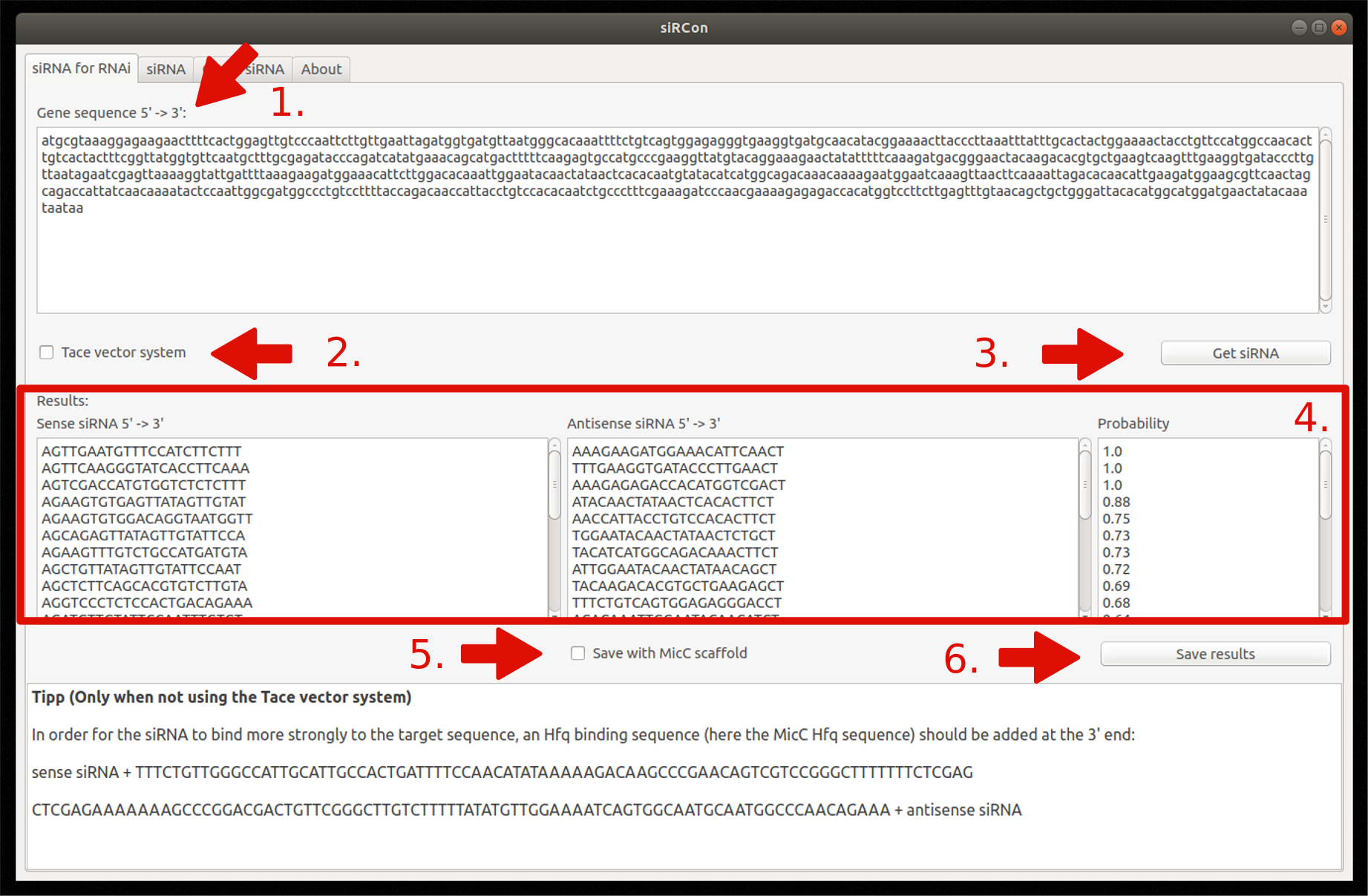
In the graphical interface, the modules are divided into different tabs (Figure 6). The last tab contains usage and copyright information.

1. siRNA for RNAi
- Insert gene sequence
- Choose Tace vector system (optionally)
- Constructions of siRNAs
- View resulting siRNAs (sense and antisense sequence) and their corresponding probability
- Decide if siRNAs should be saved with MicC scaffold (only if Tace is not used)
- Save results as FASTA file
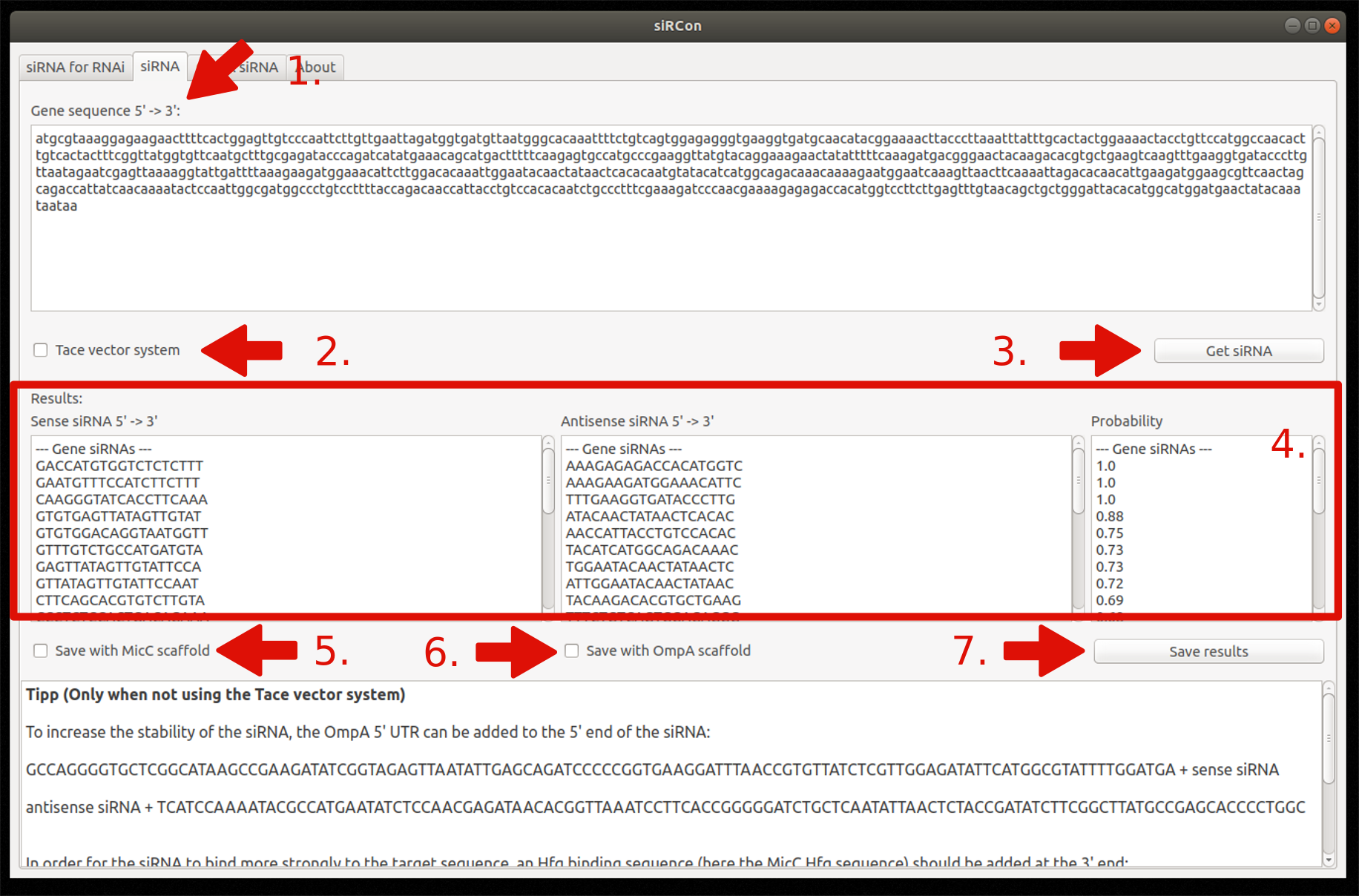
2. siRNA for silencing
- Insert gene sequence
- Choose Tace vector system (optionally)
- Constructions of siRNAs
- View resulting siRNAs (sense and antisense sequence) and their corresponding probability
- Decide if siRNAs should be saved with MicC scaffold (only if Tace is not used)
- Decide if siRNAs should be saved with OmpA scaffold (only if Tace is not used)
- Save results as FASTA file
3. Check siRNA
- Insert gene sequence
- Insert siRNA sequences
- Choose method the siRNA was constructed for (siRNA for RNAi or siRNA for silencing)
- Choose if siRNA was constructed for Tace (optionally)
- Validation of entered siRNA for given target gene sequences
- View results
- Save results (optionally)

Outlook
Elbashir, S.M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K., and Tuschl, T. (2001). Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 411: 494–498.
Foley, P.L., Hsieh, P., Luciano, D.J., and Belasco, J.G. (2015). Specificity and evolutionary conservation of the Escherichia coli RNA pyrophosphohydrolase RppH. J. Biol. Chem. 290: 9478–9486.
Kibbe, W.A. (2007). OligoCalc: an online oligonucleotide properties calculator. Nucleic Acids Res 35: W43–W46.
Na, D., Yoo, S.M., Chung, H., Park, H., Park, J.H., and Lee, S.Y. (2013). Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat. Biotechnol. 31: 170–174.
Naito, Y. and Ui-Tei, K. (2012). siRNA Design Software for a Target Gene-Specific RNA Interference. Front Genet 3.
Reynolds, A., Leake, D., Boese, Q., Scaringe, S., Marshall, W.S., and Khvorova, A. (2004). Rational siRNA design for RNA interference. Nature Biotechnology 22: 326–330.
Siomi, H. and Siomi, M.C. (2009). On the road to reading the RNA-interference code. Nature 457: 396–404.
Takasaki, S. (2009). Selecting effective siRNA target sequences by using Bayes’ theorem. Computational Biology and Chemistry 33: 368–372.
Foley, P.L., Hsieh, P., Luciano, D.J., and Belasco, J.G. (2015). Specificity and evolutionary conservation of the Escherichia coli RNA pyrophosphohydrolase RppH. J. Biol. Chem. 290: 9478–9486.
Kibbe, W.A. (2007). OligoCalc: an online oligonucleotide properties calculator. Nucleic Acids Res 35: W43–W46.
Na, D., Yoo, S.M., Chung, H., Park, H., Park, J.H., and Lee, S.Y. (2013). Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat. Biotechnol. 31: 170–174.
Naito, Y. and Ui-Tei, K. (2012). siRNA Design Software for a Target Gene-Specific RNA Interference. Front Genet 3.
Reynolds, A., Leake, D., Boese, Q., Scaringe, S., Marshall, W.S., and Khvorova, A. (2004). Rational siRNA design for RNA interference. Nature Biotechnology 22: 326–330.
Siomi, H. and Siomi, M.C. (2009). On the road to reading the RNA-interference code. Nature 457: 396–404.
Takasaki, S. (2009). Selecting effective siRNA target sequences by using Bayes’ theorem. Computational Biology and Chemistry 33: 368–372.