Team:William and Mary/Model
Math Modeling
Abstract
In order to model the behavior of incoherent feedforward loop and compare to that of naïve system, we developed a heat sensitive model which illustrates the interaction between mf-Lon, m-Scarlet and C1. We then performed analysis for both IFFL and naïve system: by including noise in input, we used stochastic model to compare noise difference and variance of the output, m-Scarlet, regarding to fixed on and off time as well as parameter regions in these two systems. In addition, we made heat maps to investigate the relationship between parameters such as duration of off time and on time of the input and performance of IFFL in distinguishing different inputs compared to naïve system. Furthermore, regarding to induced degradation rate of mf-Lon to m-Scarlet, we compared different sets of heat maps (each set has its own degradation rate) to see the impact of changing induced degradation rate of mf-Lon to m-Scarlet on superiority of IFFL to naïve system. Both of them provide insights that will inform future teams how to appropriately use IFFL under certain circumstances.
Results
Zhang C, Tsoi R, Wu F and You L demonstrated that fundamental property of incoherent feedforward loop (IFFL) can be used to exhibit temporal adaptation. They further indicated this property serves as the foundation for distinguishing input with varying temporal structure. To demonstrate this important ability, we built quantitative ordinary differential equations (ODE) model and used heat map and mesh graph to examine both vector difference and noise difference between IFFL and naïve. It is shown that under most circumstance, IFFL indeed outperforms naïve system in distinguishing different structure of temporal input. Thus IFFL is capable of interpreting information in dynamic input signal more accurately.
The conclusion we reached is that IFFL works better than Naïve system when off time is short, or the ratio of on time to duration is large. When on time is fixed, and if off time is large, or ratio of on time to duration is large, input is not sustained. Then IFFL will less likely to reach threshold and it will have stair case output, which is identical as naïve system. That means IFFL loses its ability to distinguish different type of temporary input. Therefore, short off time scenario is desired for effective use of IFFL.


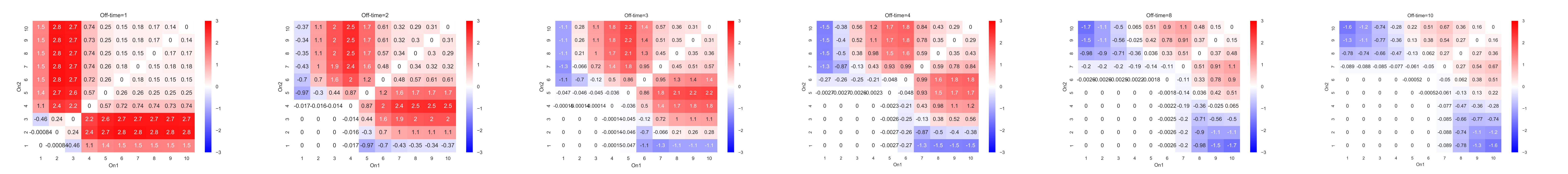
Figure 1: Vector difference between IFFL and Naïve system relative to different Off time when processing signals. We compare the vector difference between IFFL and Naïve system. vector difference is defined by the Euclidean distance of concentration expression of IFFL to Naïve at fixed off time with difference between two on time (for example, on1 = 4 and on2 = 9). When IFFL outperformed Naïve system, the vector difference is positive (red areas) and vice versa (blue areas). (Offtime1.) when off time = 1, most areas are red. So that means in this case IFFL works better in general. (Offtime10) when off time = 10, most areas are white in graph, and some parts are blue. (Bottom graph) the combined graphs of off time=1, 2, 3, 4, 8 and 10. The trend is illustrated: as off time becomes longer, IFFL gradually loses its ability to distinguish input signals.
Robustness to Noise
In Golden-Gate Assembly, type IIS restriction enzymes are used to cut DNA. Type IIS restriction enzymes are useful in that they cut outside their recognition sites, creating fragments of DNA with no unwanted base pair scars. We use the restriction enzyme BsaI, which recognizes specific DNA sequences (BsaI sites) and cuts outside of these sites, leaving sticky ends that can be ligated together with T4 DNA ligase.
In this stage of assembly, 3G takes advantage of the Cidar MoClo system, in which specific part types are distinguished by their sticky ends. After being cut, each type of part reveals a distinct sticky end on either side. The standard parts used in most synthetic circuits are promoters, 5’ untranslated regions, coding sequences, and terminators. Their MoClo sticky ends are shown in the image below:

Figure 2: to do
The unique sticky ends allow for the parts to line up in the correct sequence before being ligated together. In this way, a full transcriptional unit can be created. To prepare for the Gibson step of 3G, unique nucleotide sequences (UNS) are attached to both ends of the transcriptional unit. The UNS on the 5’ end of the transcriptional unit must have a sticky end A so that it can anneal to the promoter’s sticky end. The UNS on the 3’ end has a sticky end E so that it can anneal to the terminator’s sticky end. These sequences serve as a landing pad for primers in the next stage of PCR. They will also be used when combining the transcriptional units on to a backbone in the final stage of 3G assembly.
There are multiple 5’ UNSs and multiple 3’ UNSs, denoted by numbers (ex: UNS 1, UNS 3, UNS 10). This allows us to combine multiple fragments in the Gibson step.

Figure 3: to do
Amplification
Next, Golden-Gate products are amplified by PCR. The forward primer will anneal to the 5’ UNS and the reverse primer will anneal to the 3’ UNS. The PCR products are then gel extracted.
Gibson Stage
In the final step, a Gibson assembly is performed. Here, multiple transcriptional units are combined on to a backbone. The backbone must have the UNS 1 sequence on the 5’ end and the UNS 10 sequence on the 3’ end. This can easily be achieved by PCRing your vector with primers that add the the UNSs.

Figure 4: to do
In Gibson assembly, areas of homology are connected to form a circular plasmid. An exonuclease chews at the 5’ end of every fragment, creating compatible sticky ends that allow DNA fragments to align. DNA polymerase fills in the gaps and DNA ligase seals the bases together.
In 3G, the homologous regions between the vector and the transcriptional unit are the UNSs. To assemble a circuit with multiple transcriptional units, the first fragment should have UNS 1 on its 5’ end and the last fragment should have UNS 10 on its 3’ end. Adjacent fragments should have overlapping UNSs. For instance, the first transcriptional unit may begin with UNS 1 and end with UNS 3. The second transcriptional unit then begins with UNS 3 and ends with UNS 10. The UNS 1 on the first transcriptional unit will anneal to the UNS 1 on the vector. The UNS 3 sequences will anneal to each other, connecting the two units. Finally, the UNS 10 on the second unit will anneal to the UNS 10 on the vector, creating a circular plasmid. Of course, these steps are actually happening simultaneously, but they are outlined in this way for clarity.

Figure 5: to do
Advantages
Modularity
3G assembly offers a variety of benefits that are specifically useful for iGEM teams. This system allows for remarkable modularity at multiple levels. During the Golden Gate stage, different variants of the same part can be added to the tube to produce a variety of transcriptional units. For example, if you do not know how strong an RBS will make your circuit perform optimally, you can try several types in one reaction, then screen your circuits for the output you want. The best performing circuit can then be sequenced.

Figure 6: to do
The modularity of 3G assembly can also be seen at the Gibson stage, during which different transcriptional units can be easily combined. Once a transcriptional unit has been made, it can be combined with any other transcriptional unit with compatible UNS sequences. So, a unit flanked with UNS 1 and UNS 3 can be combined with any unit flanked with UNS 3 and UNS 5, and additionally any unit flanked with UNS 5 and UNS 10. Together, these units can go on any UNS compatible backbone. Up to six transcriptional units can be combined on to a vector at once.
Speed
The entire process of 3G assembly takes only about eight hours (including down time) followed by an overnight transformation. Furthermore, as explained above, numerous circuit variants can be created at once. Therefore, 3G is significantly more efficient than hierarchical cloning methods like ordinary Golden Gate.
Cost
Because standard adapter sequences are used, there is no need to redesign primers every time you create a circuit. This substantially cuts the cost of primers associated with Gibson assembly. In addition, the reactions in 3G are very small in volume, which saves on reagent costs. Our collaborators at UVA specifically cited this as an advantage they appreciated.
Simplicity
3G is a simple procedure that can be carried out with ease by any molecular biology lab. To demonstrate this as well as to ensure that our protocol was clear and repeatable, we collaborated with University of Pittsburgh and UVA. We sent these teams parts to assemble and test. Their success can be seen in the following gel images, which show Golden Gate PCR products of the expected lengths:

Figure 7: to do

Figure 8: to do
In addition, UVA conducted tests on a circuit they constructed. As expected, fluorescence levels were higher in the circuit with the stronger RBS. Each dot represents a biological replicate and the line is the geometric means.

Figure 9: to do
Because our collaborators were able to carry out 3G assembly, we expect that other iGEM teams should be able to as well. Thus, 3G can be practically implemented in iGEM labs, enabling teams to rapidly and inexpensively produce countless circuit variants. This leaves more time and resources for experimental work. In effect, 3G will allow for an advanced rate of scientific advances within iGEM and SynBio in general.
Within Our Project
Part One: Pad Sequences on Basic Parts
One of our first goals was to create a library of parts that can easily be used in 3G assembly. To begin this process, we designed two sequences: WM_Pad1 and WM_Pad2. These pad sequences are orthogonal to the genome of E. coli and to the UNS sequences used later in 3G assembly. We designed primers that add the pad sequences to our parts outside of the BsaI sites and inside the BioBrick Prefix and Suffix. This leads to a part that looks like the following:

Figure 10: to do
The sticky ends will, of course, vary, depending on which part we are working with. However, the pad sequences will remain the same. We put the same pad sequences on the high copy vector we commonly work with: 1C3. Then, we can easily use Gibson assembly to put our parts on the backbone and clone them. We created (number) basic parts through this method. These parts can now be used by any team for 3G assembly, and can be viewed here: link.
Part Two: General Cloning
Whenever our team needed to construct multipart circuits, we used 3G assembly. Our results were generally excellent. We designed and created 45 circuits, only some of which we are submitting to the registry. An example of a transcriptional unit we created can be seen on this gel:

Figure 11: to do
Part Three: Mixed Assembly
Our team’s project, like many iGEM projects, required us to create circuits with very specific desired behaviors. However, we were unable to perfectly anticipate what parameters would lead to the behavior we were looking for. Utilizing the modularity of 3G assembly, we took multiple variants of each part and combined them during the Golden Gate stage. This created an enormous number of transcriptional units that could be tested once put on circuits. We then screened for circuits that operated in the way we were looking for. (Link to experiment)

Figure 11: to do
Overall, 3G assembly has provided boundless opportunities for our lab, and we are excited to share this phenomenal tool with the iGEM community.