- toxicity modeling
- reactor modeling
- siRNA modeling
- ferritin structure modeling
Toxicity modeling
The toxicity of copper ions on the cell is well characterized (Ning et al., 2015) but for important parts of this project like the crossflow reactor we needed to know the exact point of time where our cells will die to achieve highest possible yields of copper. The residence time should not exceed the rate of dying and cell lysis in the system. If cell lysis kicks in copper gets released back again into the substrate media and the yield minimizes. The modeling started with the copper uptake in our cells which contains the BioBrick BBa_K2638204, which expresses oprC under pAraBAD (BBa_I0500)) control and induction at 1.0 % arabinose in. The toxicity is calculated for a single cell.The first step was to calculate the rate of expression of oprC. Therefore the characterization of BBa_I0500) of iGEM Groenningen 2011 was used to calculate the expression speed. At 1.0 % arabinose induction a raise of fluorescence of approx. ΔF = 82,000 within of t = 36,000 s was measurable. The conversion from fluorescence units to concentration in mol/L was calculated as k = 2.5 ∙ 10-6 mol/L (Furtado and Henry, 2002) and the volume of the used capillaries was V = 3.14 ∙ 10-9 L The rate of protein expression with 1.0 % arabinose is:
(3.14 ∙ 10-9 L ∙ 2.5 ∙ 10-6 mol/L ∙ 87,500) / 36,000 s = 7.85 ∙ 10-14 mol/s (1)
According to (1) the increase of OprC with t0 = 0 can be described with (2) and (3):
dOprC/dt = 7.85 ∙ 10-14 mol/s (2)
OprCt = OprC0 + t ∙ 7.85 ∙ 10-14 mol/s (3) The next step is to determine the increase of the copper concentration inside the cell. OprC transports copper ions with a rate of vmax = 0.144 mol/s into the cell and has a substrate affinity of KM = 1.66 mM (Parmar et al., 2018). Due to the missing catalytic interchange number kcat the other definition of the Michaelis-Menten equation had to be used: d(Cinside)/dt = -d(Coutside)/dt = vmax ∙ Cu2+ioutside/( Cu2+outside + KM) (4) After solving the differential equation (4) with the conditions t0 = 0 :
[Ct]- KM ∙ ln[Ct] = vmax ∙ t + [C0] - KM ∙ ln [C0] (5) Due to the fact that this only describes the kinetic of a single molecule of OprC the (3) and the Avogadro number NA = 6.022 ∙ 1023 mol-1 . That way the addition was possible of the both terms on the right side of (5) and the end equation results in:
[Ct]- KM ∙ ln[Ct] = vmax ∙ t2 ∙ NA ∙ 7.85 ∙ 10-14 mol/s + [C0] - KM ∙ ln [C0] (6) At this point the equation is not further to simplify. Because of this a short python script trying out multiple different number constellations had to be tried out.
kurve = []
for each in listCt[:end+1]:
for t in ts:
nOPRC = t*5*(10**(-29))
#print each
#print t
gleichung1 = vmax*t*nOPRC*6.022*(10**(23)) - C0 - math.log(C0)*KM
gleichung2 = -each - KM * math.log(each)
gleichung2small = gleichung2 - gleichung2*0.00001
gleichung2big = gleichung2 + gleichung2*0.00001
if gleichung2small <= gleichung1 and gleichung2big >= gleichung1:
kurve.append((each,t))
print each
print t
print kurve
with open ("Tupel_fuer_Kurve3.txt", "w") as out:
This way the toxic copper concentration ctox = 1 ∙ 10-5 mol/L (Ning, 2015) after inside the cell is approx. reached after the induction with 1.0 % arabinose.
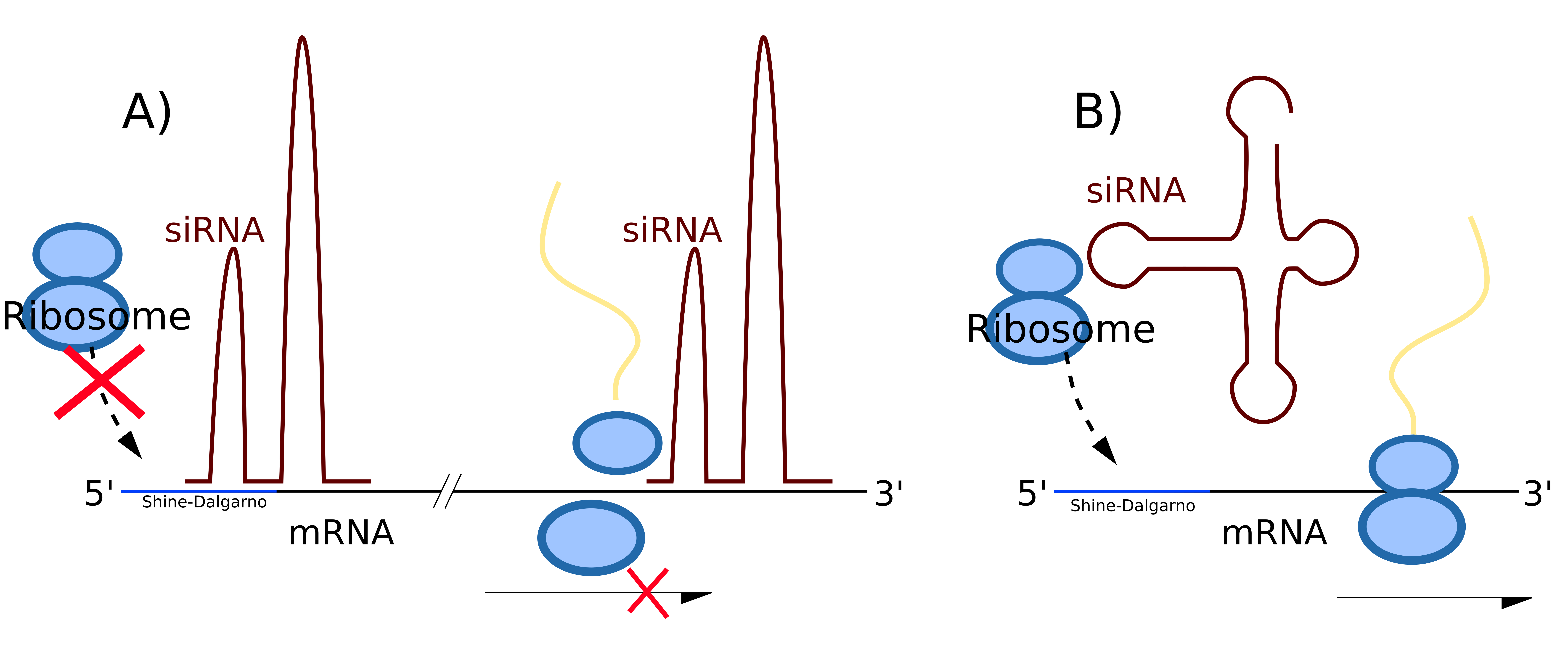
siRNA promoter model
First the most fitting siRNAs for our project has to be calculated. A program will be written which returns a selection of compatible siRNAs for a gene you want to silence. Possible secondary structures should also be considered (Mathews, 1999) because they could prevent the silencing mechanism of siRNAs.