Team:William and Mary/Results
Results

Construction of an Abstract IFFL Model:
The first step in designing our decoding circuit was to examine what circuit architectures were capable of distinguishing between different types of dynamical inputs. After reviewing the literature, we found that many natural systems that utilize dynamic information encoding share a common circuit motif, that of the incoherent feedforward loop (IFFL). The IFFL is an architecture in which an input activates the production of both a reporter/output as well the production of an inhibitor of that reporter (Figure 1) [1]. Furthermore, this IFFL architecture has been shown to be capable of serving as a temporal distinguisher, meaning it can differentiate between different types of temporal (dynamic) signals.

Figure 1: Schematic of an Incoherent Feed Forward Loop architecture. An activator (green) activates the production of a reporter/output (purple) as well as an inhibitor of the reporter (blue).
Based on our understanding that incoherent feedforward loops (IFFLs) could serve as temporal distinguishers, the first goal of our project was to replicate the model from Zhang, et al. (2016) [1]. We constructed an abstract kinetics based IFFL model, and found that IFFLs are indeed temporal classifiers. We found that when an IFFL was given an oscillatory input signal it will give qualitatively different outputs depending on the length of input signals (Figure 2). In the presence of long input signals, an IFFL will give output pulses of a fixed height, whereas in the presence of shorter input signals it will produce a staircase like output where the output concentration increases with each new input pulse.

Figure 2: The results from our abstract IFFL model. When inputs are long (left), reporter concentration pulses at a fixed height in response to each input. When input lengths are shorter than the threshold time, each input pulse will generate a stepwise increase whose size is determined by pulse time.
The ultimate source of this behavior is the dynamics of the inhibitor; when inputs are short, the amount of inhibitor never crosses the threshold at which it efficiently degrades the output, leading to stepwise increases in the output. Conversely, when inputs are long, the inhibitor crosses the threshold and reduces the concentration of the output. Importantly for our project, these results showed that IFFLs are capable of distinguishing the temporal properties of inputs, giving different outputs depending on the length of the input signal. That means that an IFFL can provide the basis for a system that is attempting to interpret dynamic, i.e. time encoded information.
Creating a Protease Based IFFL:
With the knowledge that an IFFL would provide a suitable architecture for the construction of our decoder, we decided to use a previously characterized IFFL system that utilizes the mf-Lon protease system [2,3]. In this system the mf-Lon protease acts as an inhibitor by degrading a reporter with a protein degradation tag (pdt) (Figure 3).

Figure 3: Schematic of the dual inducible mScarlet-I-pdt mf-Lon IFFL system. Inducers act as independently tunable proxies for inputs and mf-Lon acts as an inhibitor of the reporter mScarlet-I pdt.
This is an extremely powerful and tunable system because the degradation rate constant can be tuned by using different strength pdts. By simultaneously activating the production of mf-Lon and mScarlet-I-pdt with two separate small molecule inducers, this system operates as an IFFL, with the induction of the circuit leading to the production of both the output (mScarlet-I-pdt) and its inhibitor (mf-Lon). However, by using two separate inducible systems, the production rate of the inhibitor and the output can be tuned independently of one another. Last year, work in iGEM confirmed that this system is capable of generating a pulsatile output from a continuous signal. Our first experiment therefore was to test if we could replicate this behavior on the plate reader, which would serve the dual purpose of confirming one half of the theoretical basis of our model as well as allow us to comfortably use the plate reader for future experiments involving multiple conditions. Using the registry parts K2333434 (plLac mf-Lon) and K2333428 (ptet mScarlet-I pdt#3) we were able to successfully replicate this pulsatile behavior given a constant input (Figure 4).

Figure 4: Normalized to max fluorescence of the chemically induced circuits BBa_K2333427 and BBa_K2333434 when grown at 37C and then chemically induced with 50ng/mL ATC and 0.1mM IPTG. Dots represent the geometric mean of 3 distinct biological replicates (colonies) and the blue shaded region represents one geometric standard deviation above and below the mean.
Testing Short Inputs:
We next wanted to confirm that this system would be capable of discerning between temporal inputs of different lengths. However, in order to do this we would first need to be able to determine how we could dynamically change the concentration of inducer present in solution. While our first choice would have been to use a constantly flowing microfluidics chamber along with a fluorescence microscope, we were unable to do so because of the cost involved and lack of institutional experience with such devices. Instead, we developed our own protocol in which centrifugation was used to pellet cells, allowing us to wash them and replace the media with fresh noninducer-containing media. However, we had concerns with whether this protocol would be able to remove inducer without otherwise impacting the results of experiment. One obvious concern was that since spinning down cells takes a significant amount of time, the precision of our experiments could be compromised, given that they involve the precise timing of activation periods. Other potential issues included the impact that variable temperature and the forces of centrifugation could have on gene expression.
Because of these potential pitfalls, we recognized that our first step had to be confirming that this method is valid. In our first few trials, we had issues choosing the proper speed for the centrifuge and found that our cell density was too low to form substantial cell pellets. However, with time, we were able to improve upon our procedure, eventually coming to a procedure wherein we centrifuged 50 mL of cells at 2762 RCF for 3 minutes, washed with prewarmed media, and then repeated and resuspended cells. When we tested if this method was effective at controlling the expression of a pTet inducible mScarlet by removing ATc, we found that it was effective. (Figure 5).

Figure 5: Fluorescence/OD600 (AU) measurements of the chemically induced circuits BBa_K2333427 and BBa_K2333434 when grown at 37C and then chemically induced with 100ng/mL ATC and 0.1mM IPTG from times 0-120. At timepoint 120, each replicate was split into two tubes and spun down for three minutes at 2762 RCF. Half of the cells were washed with media without ATC, and the other half were washed with media containing ATC. Dots represent the geometric mean of 3 distinct biological replicates (colonies) and the blue shaded region represents one geometric standard deviation above and below the mean.
With a way to dynamically alter the input of our circuits, we next wanted to confirm the prediction that short inputs would lead to a stepwise increase in output concentration. We tested this prediction in the small molecule induced IFFL by taking single cell fluorescence measurements of an input paradigm of 40 minutes on, 40 minutes off, 40 minutes on. Using this paradigm we were able to validate the prediction of the model, showing that IFFLs do indeed function as temporal distinguishers (Figure 6).

Figure 6: Normalized (% max) fluorescence measurements of the chemically induced circuits BBa_K2333429 (pTet mScarlet-I pdt) and BBa_K2333434 (plLac mf-Lon) when grown at 37C and then chemically induced with 100ng/mL ATC and 0.1mM IPTG from times 0-40 and 80-120 (represented by shaded region). Dots represent the geometric mean of at least calibrated 10,000 single cell measurements of 3 distinct biological replicates (colonies) and the blue shaded region represents one geometric standard deviation above and below the mean.
However, when we tried to extend our results we had trouble testing more cycles of inputs and shorter input pulse durations. Often times after the second centrifugation step we would obtain a small or nonexistent size pellet of cells, resulting in us have to abort the experiment. While we considered that it might be possible to scale up the size of our culture volumes, we reasoned that for this reason as well as others, the use of the chemically inducible system would limit our ability to characterize interesting dynamic patterns of input. For more information on our tests with this system, see the chemically inducible system page.
We evaluated a number of potential types of induction before settling on using the temperature sensitive CI (ts-CI) system. Temperature based induction offers a variety of unique benefits. First, temperature is easy to control in using standard lab equipment such as incubators and plate readers, so we would not require any equipment we don’t already have. Secondly, temperature is relevant in many in vivo systems such as those that regulate the pathogenicity of bacteria [4], meaning that our synthetic system would be potentially useful for future groups working in that area.
Modeling a Temperature Sensitive Decoding IFFL:
Having decided upon our new system, we next wanted to create a more biologically based mathematical model to evaluate the decoding properties of this system in particular. To do so, we developed an ODE model of our ts-CI system. We determined that our system functions as an IFFL and then sought to investigate how well our decoding circuit was able to classify different types of dynamic signals.
We first investigated how well our decoding circuit was able to distinguish between different length oscillatory inputs. Comparing the outputs of both our decoding circuit and a naive tracker (i.e. a reporter which is activated the input), we were able to determine that the decoder was superior at discerning the differences between different temporal inputs (Figure 7). Moreover, we found that the decoding circuit was particularly good at discerning even small differences in temporal inputs.

Figure 7: Left. Heat map comparing the euclidian distance of outputs for a given change in input pulse duration. Redder values indicate that the decoder is better able to discern the difference between the inputs. Right. Graphs showing the behavior of the input (top), IFFL (2nd from the top), naive model (3rd from the top), and the difference between each model for a specific change in input pulse durations (bottom). Larger differences indicate higher temporal discrimination.
Given that the decoding circuit is so sensitive to small changes in input, it would be reasonable to assume that it is also more vulnerable to noisy inputs. To test how susceptible the decoder was to noisy inputs, we added gaussian noise to the model inputs and tested the resulting output variance for a variety of different parameter regions. Surprisingly, we found that in fact the decoder has a lower output variance than the naive model when given noisy inputs (Figure 8). This is due to the proportionality effect of mf-Lon, which causes stochastic variations in the input to be weakened, leading to a less variable output. In total, our modeling shows that the decoding circuit is capable of discerning even very small temporal inputs, while still performing exponentially better in response to input variability when compared to a naive system. This implies that our system and other IFFL decoding systems represent a solution to the problem of interpreting dynamic and noisy data. To see more about how we explored the ways in which an IFFL can be made to perform superiorly to a naive circuit see our modeling page.
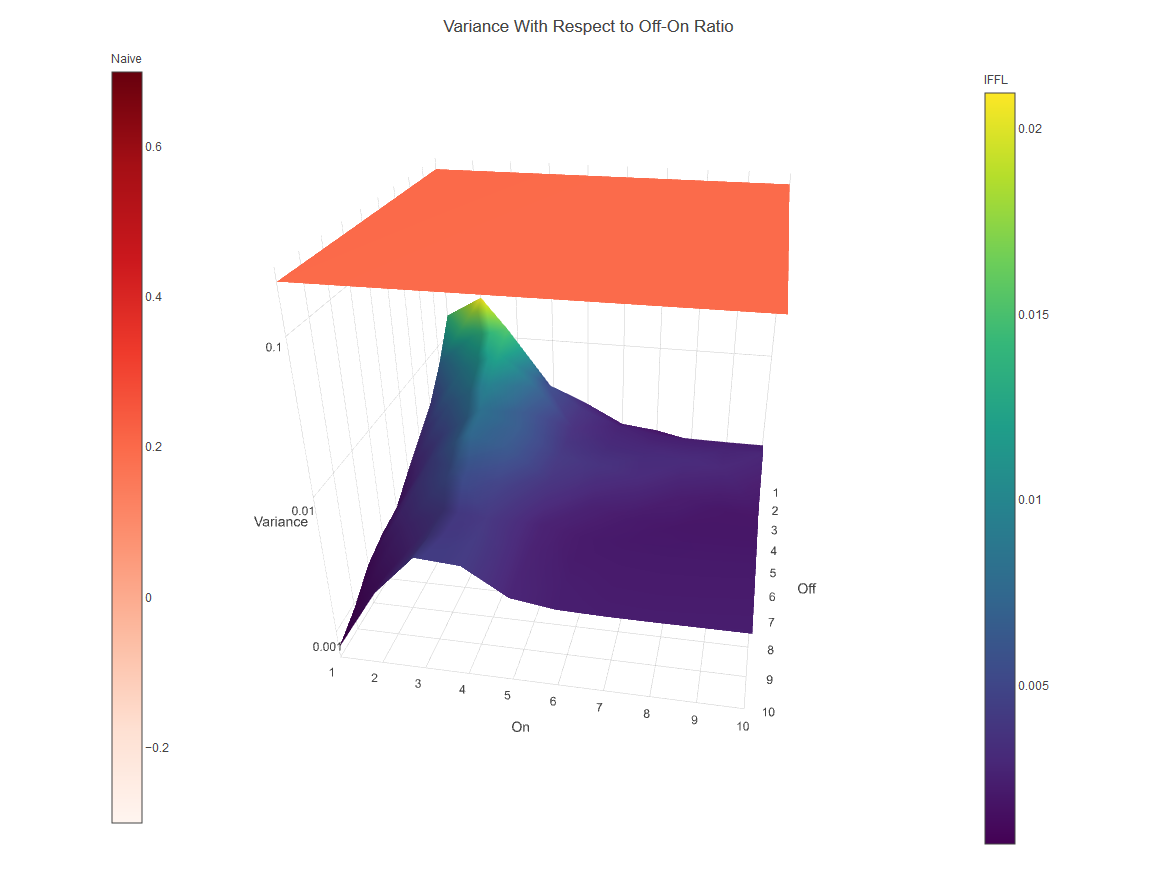
Figure 8: Input variance as direct translation to noise. Variance of IFFL with respect to all combination of off and on time is exponentially lower than that of Naive.
Implementation of a Temperature Sensitive IFFL:
Having confirmed that a heat sensitive decoder would perform better than a naive tracker, and validated that the original model’s predictions, we next attempted to put our heat sensitive IFFL into practice. We designed and constructed a number of heat sensitive IFFL systems using a ts-CI repressible system with 3G Assembly. During this testing we were able to validate that our parts were functional, yet we were unable to find a combination of parts with appropriate parameters to create a decoding IFFL. In each case, the IFFL circuits we created lead to either no degradation of reporter (a too high reporter/mf-Lon ratio), no fluorescence (a too high mf-Lon/reporter ratio) or cell death (too high mf-Lon).
We attempted to solve this problem in a number of different ways. We tried combining our circuits onto a single plasmid construct, genomically integrating constructs, and adding a strong degradation tag to mf-Lon. However, none of those approaches were successful in obtaining a temperature sensitive IFFL with the appropriate parameters. We next decided to shift to an alternative temperature sensitive system utilizing the temperature sensitive repressor TlpA and its cognate promoter operators. However, despite many iterative design build test cycles, we were unable to get any of these approaches to produce IFFLs which outputted the characteristic pulse when activated. See our page on temperature inducible systems.
Moving forward, we believe that chemically induced systems show the most promise. Our preliminary data indicates that with the proper equipment (microfluidics), chemically inducible IFFLs may serve as a tool to distinguish temporal inputs. In addition to chemical induction, however, there is potential for other types of inducible IFFLs to act as decoders, including optogenetic systems and other temperature sensitive systems such as those utilizing riboswitches or RNA thermometers.
References
[1] Zhang C, Tsoi R, Wu F, You L (2016) Processing Oscillatory Signals by Incoherent Feedforward Loops. PLoS Comput Biol 12(9): e1005101. doi:10.1371/journal.pcbi.1005101
[2] D Ewen Cameron and James J Collins. Tunable protein degradation in bacteria. Nature biotechnology, 32(12):1276–1281, 20
[3] https://2017.igem.org/Team:William_and_Mary/Dynamic_Control
[4] Gal-Mor O, Valdez Y, Finlay BB. The temperature-sensing protein TlpA is repressed by PhoP and dispensable for virulence of Salmonella enterica serovar Typhimurium in mice. Microbes Infect / Institut Pasteur. 2006;8:2154–62. doi:10.1016/j.micinf.2006.04.015.