

Short Summary
- toxicity modeling
- reactor modeling
- siRNA modeling
- ferritin structure modeling
Toxicity modeling
The toxicity of copper ions on the cell is well characterized (Ning et al., 2015) but for important parts of this project like the crossflow reactor we needed to know the exact point of time when our cells will die to achieve highest possible yields of copper.
The residence time should not exceed the rate of dying and cell lysis in the system. If cell lysis kicks in copper gets released back again into the substrate media and the yield minimizes. The modeling started with the copper uptake in our cells containing the BioBrick BBa_K2638204, which expresses oprC under pAraBAD (BBa_I0500)) control and induction at 1.0 % arabinose in. The toxicity is calculated for a single cell.
The first step was to calculate the rate of expression of oprC. Therefore the characterization of BBa_I0500) of Groningen was used to calculate the expression speed. At 1.0 % arabinose induction a raise of fluorescence of approx. ΔF = 82,000 within of t = 36,000 s was measurable.
The conversion from fluorescence units to concentration in mol/L was calculated as k = 2.5 * 10-6 mol/L (Furtado and Henry, 2002) and the volume of the used capillaries was V = 3.14 * 10-9 L The rate of protein expression with 1.0 % arabinose is:
$$\frac{3.14 * 10^{-9} L * 2.5 * 10^{-6} mol L^{-1} * 87,500}{36,000 s} = 7.85 * 10^{-14} mol*s^{-1} (1)$$
can be described with (2) and (3): $$\frac{dOprC}{dt} = 7.85 * 10-14 mol*s^{-1}$$ (2) $$OprC_t = OprC_0 + t ∙ 7.85 * 10^{-14} mol*s^{-1}$$ (3)
kurve = []
for each in listCt[:end+1]:
for t in ts:
nOPRC = t*5*(10**(-29))
#print each
#print t
gleichung1 = vmax*t*nOPRC*6.022*(10**(23)) - C0 - math.log(C0)*KM
gleichung2 = -each - KM * math.log(each)
gleichung2small = gleichung2 - gleichung2*0.00001
gleichung2big = gleichung2 + gleichung2*0.00001
if gleichung2small <= gleichung1 and gleichung2big >= gleichung1:
kurve.append((each,t))
print each
print t
print kurve
with open ("Tupel_fuer_Kurve3.txt", "w") as out:

siRNA promoter model
First the most fitting siRNAs for our project has to be calculated. A program will be written which returns a selection of compatible siRNAs for a gene you want to silence. Possible secondary structures should also be considered (Mathews, 1999) because they could prevent the silencing mechanism of siRNAs.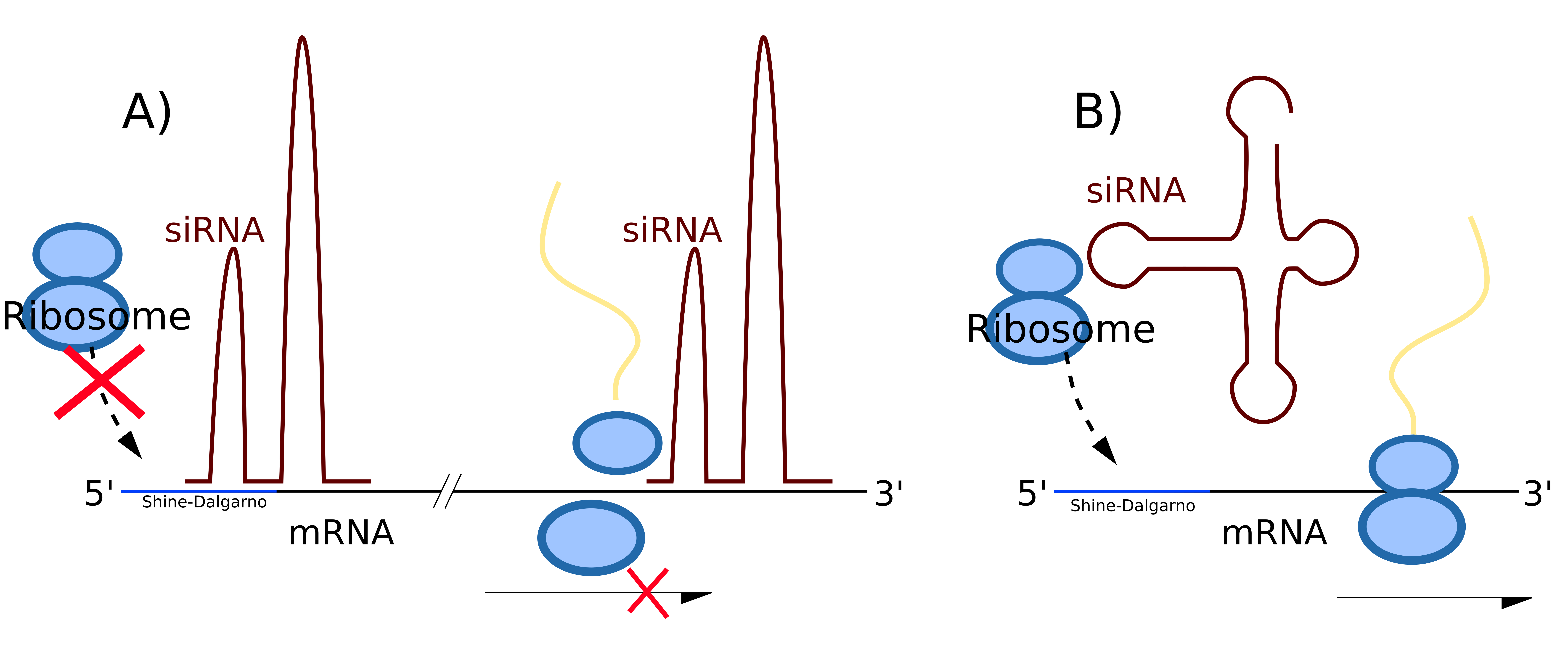
Discussion
The modeling for the point of time where the toxicity kicks in showed, that due to the troublesome way to find the right promoter strength a universal promoter library would be an important addition to the synthetic biology community. That way the promoter library in the parts collection represents an universal standard for the iGEM community to characterize their promoters.The crossflow reactor design and modeling was strongly influenced by the toxicity modeling.Another way to approach this problem were the BioBricks BBa_K2638100, BBa_K2638101, BBa_K2638103 BBa_K2638105, BBa_K2638106, BBa_K2638120 and BBa_K2638121 enrich now the parts registry.
The siRNA/RNAi modeling went over into our software project because it showed way more potential like this. The ferritin modeling was a important measure to make sure the characterization of the ferritin molecules didn’t encountered any issues.
References
Barnes, D. J., & Chu, D. (2010). Introduction to modeling for biosciences. Springer Science & Business Media.
Delihas, N., & Forst, S. (2001). MicF: an antisense RNA gene involved in response of Escherichia coli to global stress factors. Journal of molecular biology, 313(1), 1-12.
Furtado, A., & Henry, R. (2002). Measurement of green fluorescent protein concentration in single cells by image analysis. Analytical biochemistry, 310(1), 84-92. Ning, C., Wang, X., Li, L., Zhu, Y., Li, M., Yu, P., ... & Zhang, Y. (2015). Concentration ranges of antibacterial cations for showing the highest antibacterial efficacy but the least cytotoxicity against mammalian cells: implications for a new antibacterial mechanism. Chemical research in toxicology, 28(9), 1815-1822.
Mathews, D. H., Sabina, J., Zuker, M., & Turner, D. H. (1999). Expanded sequence dependence of thermodynamic parameters improves prediction of RNA secondary structure. Journal of molecular biology, 288(5), 911-940.
Parmar, J. H., Quintana, J., Ramírez, D., Laubenbacher, R., Argüello, J. M., & Mendes, P. (2018). An important role for periplasmic storage in Pseudomonas aeruginosa copper homeostasis revealed by a combined experimental and computational modeling study. Molecular microbiology.
Phan, C. M., & Nguyen, H. M. (2017). Role of capping agent in wet synthesis of nanoparticles. The Journal of Physical Chemistry A, 121(17), 3213-3219.
Stach, J. E., & Good, L. (2011). Synthetic RNA silencing in bacteria–antimicrobial discovery and resistance breaking. Frontiers in microbiology, 2, 185.
Delihas, N., & Forst, S. (2001). MicF: an antisense RNA gene involved in response of Escherichia coli to global stress factors. Journal of molecular biology, 313(1), 1-12.
Furtado, A., & Henry, R. (2002). Measurement of green fluorescent protein concentration in single cells by image analysis. Analytical biochemistry, 310(1), 84-92. Ning, C., Wang, X., Li, L., Zhu, Y., Li, M., Yu, P., ... & Zhang, Y. (2015). Concentration ranges of antibacterial cations for showing the highest antibacterial efficacy but the least cytotoxicity against mammalian cells: implications for a new antibacterial mechanism. Chemical research in toxicology, 28(9), 1815-1822.
Mathews, D. H., Sabina, J., Zuker, M., & Turner, D. H. (1999). Expanded sequence dependence of thermodynamic parameters improves prediction of RNA secondary structure. Journal of molecular biology, 288(5), 911-940.
Parmar, J. H., Quintana, J., Ramírez, D., Laubenbacher, R., Argüello, J. M., & Mendes, P. (2018). An important role for periplasmic storage in Pseudomonas aeruginosa copper homeostasis revealed by a combined experimental and computational modeling study. Molecular microbiology.
Phan, C. M., & Nguyen, H. M. (2017). Role of capping agent in wet synthesis of nanoparticles. The Journal of Physical Chemistry A, 121(17), 3213-3219.
Stach, J. E., & Good, L. (2011). Synthetic RNA silencing in bacteria–antimicrobial discovery and resistance breaking. Frontiers in microbiology, 2, 185.