Difference between revisions of "Team:TJU China/Model"
| Line 536: | Line 536: | ||
</div> | </div> | ||
| − | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/d/df/T--TJU_China--z4.png"></div> | + | <div class="pic"> |
| + | <img src="https://static.igem.org/mediawiki/2018/d/df/T--TJU_China--z4.png"> | ||
| + | </div> | ||
<div class="figure">Figure 4</div> | <div class="figure">Figure 4</div> | ||
| − | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/f/f7/T--TJU_China--z5.png"></div> | + | <div class="pic"> |
| + | <img src="https://static.igem.org/mediawiki/2018/f/f7/T--TJU_China--z5.png"> | ||
| + | </div> | ||
<div class="figure">Figure 5</div> | <div class="figure">Figure 5</div> | ||
<div class="word">Where k is the reaction rate constant; f represents the forward reactions;b represents the backward reaction.And </div> | <div class="word">Where k is the reaction rate constant; f represents the forward reactions;b represents the backward reaction.And </div> | ||
| − | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/e/ec/T--TJU_China--zm1.PNG"></div> | + | <div class="pic"> |
| + | <img src="https://static.igem.org/mediawiki/2018/e/ec/T--TJU_China--zm1.PNG"> | ||
| + | </div> | ||
<div class="word">So for a complete match:</div> | <div class="word">So for a complete match:</div> | ||
| − | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/0/00/T--TJU_China--zm2.png"></div> | + | <div class="pic"> |
| + | <img src="https://static.igem.org/mediawiki/2018/0/00/T--TJU_China--zm2.png"> | ||
| + | </div> | ||
<div class="word">Consider the rate constant $K_f(i)$ and #k_b(i)$:</div> | <div class="word">Consider the rate constant $K_f(i)$ and #k_b(i)$:</div> | ||
| − | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/b/b2/T--TJU_China--zm3.png"></div> | + | <div class="pic"> |
| − | <div class="word">where $F_i$ means free energy of each metastable state,$T_{i,i+1}$means the highest free energy point on the reaction path from position i | + | <img src="https://static.igem.org/mediawiki/2018/b/b2/T--TJU_China--zm3.png"> |
| − | + | </div> | |
| + | <div class="word">where $F_i$ means free energy of each metastable state,$T_{i,i+1}$means the highest free energy point on the reaction | ||
| + | path from position i to position i+1.Therefore,$T_{i,i+1}$-$F_i$ is the activation energy of forward reaction and | ||
| + | $T_{i,i+1}$-$F_i$ is activation energy of the backward reaction. | ||
| + | </div> | ||
| + | <div class="pic"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/0/02/T--TJU_China--zm4.png"> | ||
</div> | </div> | ||
| − | |||
<div class="word">We define</div> | <div class="word">We define</div> | ||
| − | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/f/fe/T--TJU_China--zm5.png"></div> | + | <div class="pic"> |
| + | <img src="https://static.igem.org/mediawiki/2018/f/fe/T--TJU_China--zm5.png"> | ||
| + | </div> | ||
<div class="word">So</div> | <div class="word">So</div> | ||
| − | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/9/93/T--TJU_China--zm6.png"></div> | + | <div class="pic"> |
| − | <div class="word"></div> | + | <img src="https://static.igem.org/mediawiki/2018/9/93/T--TJU_China--zm6.png"> |
| + | </div> | ||
| + | <div class="word">From the above,it is clear that the matching probability depends only on the state transition energy,not on the free | ||
| + | energy of the metastable states.If we assume there is one dominant minimal bias,say for n = n ∗ ,then this equation | ||
| + | can be approximated as:</div> | ||
| + | |||
| + | <div class="pic"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/7/72/T--TJU_China--zm7.png"> | ||
| + | </div> | ||
| + | <div class="word"> | ||
| + | To sum up,the cleavage possibility mainly relies on the free energy change, and PAM appears as a significant energy decline. | ||
| + | </div> | ||
| + | <div class="pic"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/7/7c/T--TJU_China--AT.png"> | ||
| + | </div> | ||
| + | <div class="figure">Figure 6:AT</div> | ||
| + | <div class="pic"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/3/37/T--TJU_China--CG.png"> | ||
| + | </div> | ||
| + | <div class="figure">Figure 7:CG</div> | ||
| + | <div class="word">So the kinetic module set up a form to regress the relationship between cleav age and the numbers of nucleotide matches | ||
| + | and mismatches.In consideration of this problem more carefully,the cleavage possibility becomes equal to analysis | ||
| + | energy change,and we know the binding energy of A/T and C/G is diferent due to the different hydrogen bond between | ||
| + | them.However,in appearing kinetic mod el,research tend to describe them in a rough definition as “matched base pairs”, | ||
| + | and the energy incline in C/G is approximately 1.5 folds as A/T.Similarly,the mismatch has more difference because | ||
| + | the size of nucleotides is various.Hence, the combination of the mismatched base pair was classified by group volume,i.e. | ||
| + | two pyrimidines(such as C/T,“L”),pyrimidine and purine(such as C/A,“M”),t wo purine(such as G/T,“S”).Hence,our possibility | ||
| + | can be calculated using the following formation. | ||
| + | </div> | ||
| + | <div class="pic"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/5/53/T--TJU_China--zm8.png"> | ||
| + | </div> | ||
| + | <div class="subtitle">2.2 Optimization module</div> | ||
| + | <div class="word">It is a common sense that experimental results are facts,but theoretical results are only conjectures.From kinetic module,we | ||
| + | can get an output,which is the cleavage possibility.The parameter we choose only aims to make results have discrimination,while | ||
| + | it’s not quantitative.And in a cleavage experiment,we only have two outcomes,successful and unsuccessful.To make | ||
| + | our prediction possibility more approximate toexperiment,we regard this as a regression problem. | ||
| + | <br> Here,the method we choose is stochastic gradient descent(SGD) and cross entropy.And their principle can be concluded | ||
| + | as follows. | ||
| + | </div> | ||
| + | <div class="pic"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/a/a4/T--TJU_China--zm9.png"> | ||
| + | </div> | ||
| + | <div class="word">where θ means the parameters array and J means the loss function. Considering the difference in gradient calculation,we | ||
| + | use difference to substi tute differential aim to accelerate operating speed.</div> | ||
| + | <div class="pic"> | ||
| + | <img src="https://static.igem.org/mediawiki/2018/f/f3/T--TJU_China--zm10.png"> | ||
| + | </div> | ||
| + | <div class="word">By using this simple method,our model can be more vibrant,updating using newest data and becoming more reliable.</div> | ||
| + | |||
| + | <div class="subtitle">2.3 Pre-selector</div> | ||
| + | <div class="word">It’s obviously that the algorithm is too complex to applying in slide in a huge | ||
| + | DNA array.To solve this problem,we use a pre-selector to get some candidates | ||
| + | and use previous model to compare them so that we could get a greatest target.<br> | ||
| + | And here this pre-selector structure is very simple.Considering use this map | ||
| + | to reflect the similarity between target and full DNA.</div> | ||
| + | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/2/27/T--TJU_China--zm11.png"></div> | ||
| + | <div class="word">Here,we use PAM as an input and collect the array which contain the same | ||
| + | beginning code as PAM.</div> | ||
| + | <div class="title">3 Result</div> | ||
| + | <div class="word">Here,we set the parameters as default values and observe its performance.As | ||
| + | the following figure shows,the energy always decreases or has some turning point | ||
| + | and is always negative.Such as the red line,it has a peak due to a mismatch | ||
| + | here,and in our model,we find that it doesn’t make the energy positive.That | ||
| + | means that in this reaction process there is some force like ”momentum” pushing | ||
| + | it to proceed and cross the energy peak.Corresponding to the other figure’s two | ||
| + | particular locations(aandb),only in these points their energy are all negative | ||
| + | (because we want to see the idea target series,only the locations which correspond | ||
| + | to negative energy are collected).After testing our code runtime,its manage rate | ||
| + | can reach approximate 2×$10^8$ base/h (under parallel calculation in 4 cores) and | ||
| + | have somewhat applicaiton value.Besides the default parameters,we hope our | ||
| + | model can hit more true data.So if we get the experiment data,we can use model | ||
| + | 2.2 to get greater parameters.(@@no experiment data)</div> | ||
| + | |||
| + | <div class="pic"><img src="https://static.igem.org/mediawiki/2018/1/1c/T--TJU_China--energy_change.png"></div> | ||
| + | <div class="figure">Figure 8 energy change</div> | ||
| + | |||
| + | <div class="head">References</div> | ||
| + | <div class="word">[1] family=Klein,familyi=K.,given=Misha,giveni=M.,"Hybridization kinet | ||
| + | ics explains CRISPR-Cas off-targeting rules".In:Cell reports 22.6(2018),pp.1413-1423. | ||
| + | </div> | ||
| + | |||
| + | |||
| + | |||
| + | |||
Revision as of 01:18, 17 October 2018
<!DOCTYPE >







Dynamic Model of Heavy Metal Detection Biosensor
Minghui Yin,Sherry Dongqi Bao
TianJin University
October 15,2018
TianJin University
October 15,2018
1 Introduction
Modeling is a powerful tool in synthetic biology. It provides us with a necessary engineering approach to characterize
our pathways quantitatively and predict their performance,thus help us test and modify our design.Through the dynamic
model of heavy-metal detection biosensor,we hope to gain insights into the characteristics of our whole circuit's
dynamics.
2 Methods
2.1 Analysis of metabolic pathways
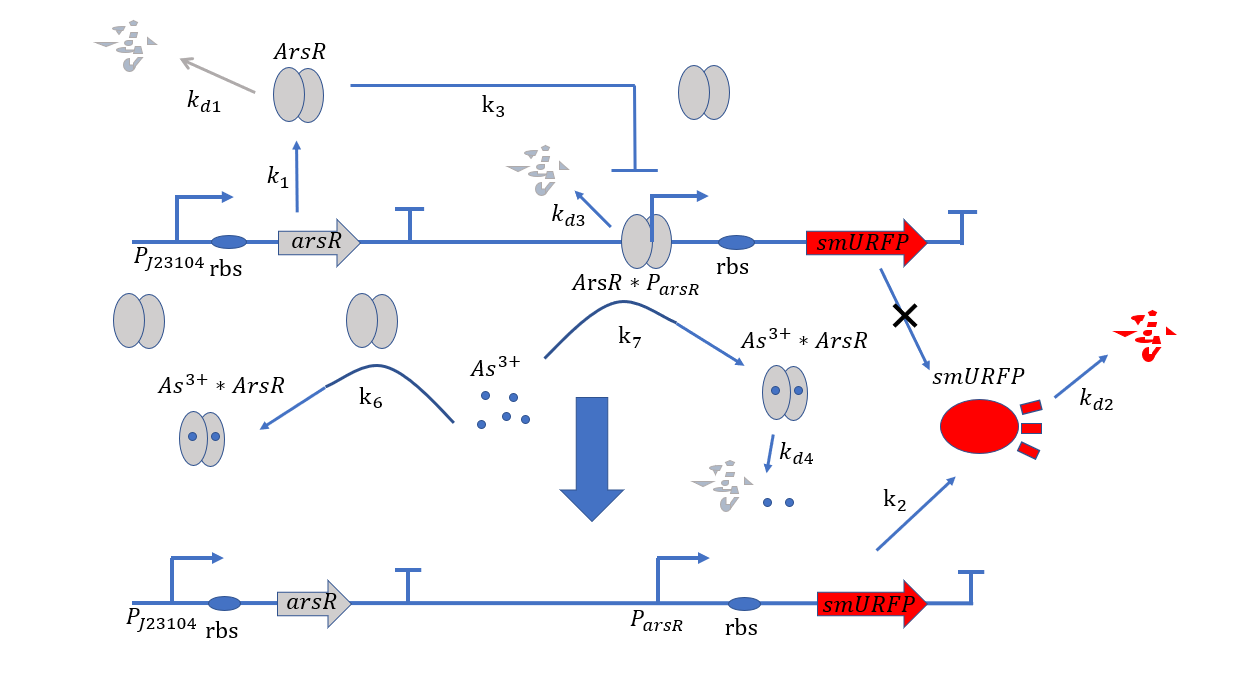
Figure 1: Metabolic pathways related to plasmid#1
At the beginning, on the plasmid#1, the promoter $P_{arsR}$ isn't bound with ArsR,thus it is active.ArsR and smURFP are
transcribed and translated under the control of the promoters $P_{arsR_{u}}$ and $P_{arsR_{d}}$,with subscript u
and d representing upstream and downstream separately.The subscript l of smURFP in the equation means leaky expression
without the expression of $As^{3+}$.As ArsR is expressed gradually,it will bind with the promoter $P_{arsR}$ and
make it inactive.[1]
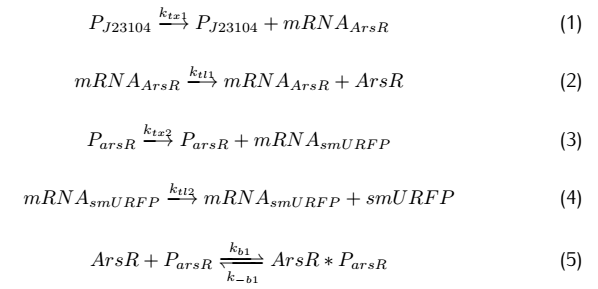
On the plasmid#2,the fusion protein of dCas9 and RNAP(RNA polymerase) are produced after transcription and translation,and
sgRNA is produced after transcription.
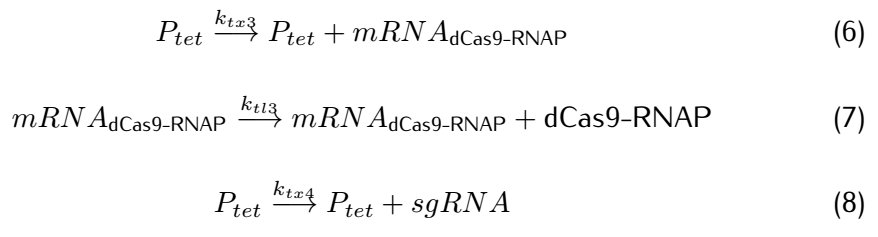
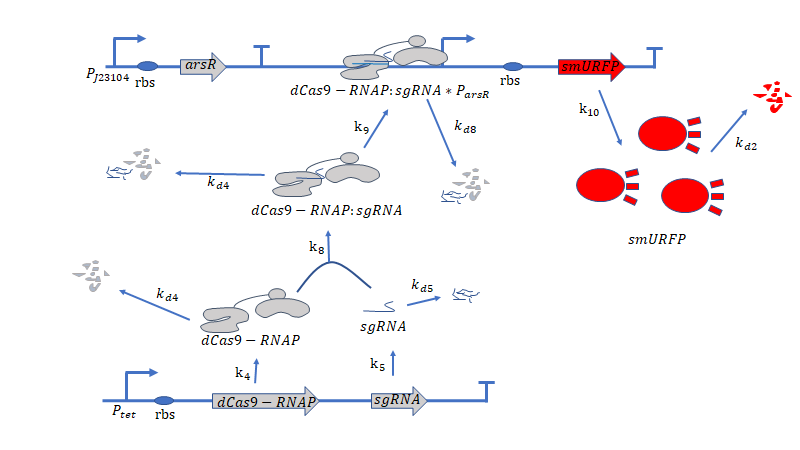
Figure 2: Metabolic pathways related to dCas9/RNAP
dCas9(*RNAP) can bind with its target DNA sequence without cutting, which is at the upstream of the promoter $P_{arsR_{d}}$.Simulataneously,dCas9
can lead RNAP to bind with the promoter $P_{arsR_{d}}$ and enhance the transcription of smURFP.However,because the
promoter $P_{arsR_{d}}$ has already bound with ArsR,as a result,RNAP can't bind with the promoter $P_{arsR_{d}}$.
can’t bind with the promoter $P_{arsR_{d}}$.
However,at the presence of $As^{3+}$,it can bind with ArsR,then dissociate ArsR and $P_{arsR_{d}}$ , which makes the
combination of RNAP and $P_{arsR_{d}}$ possible.
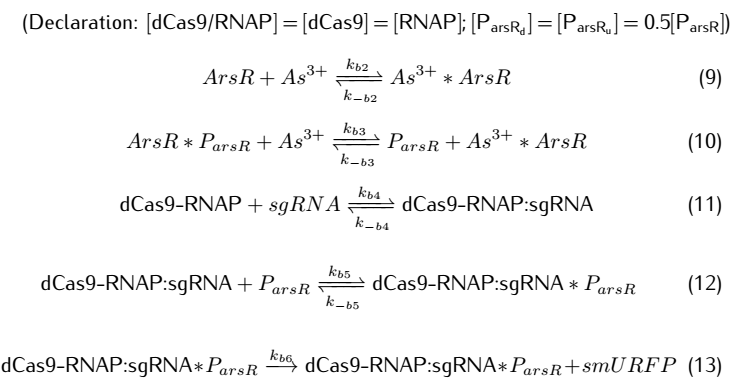
We then take degradation into account:

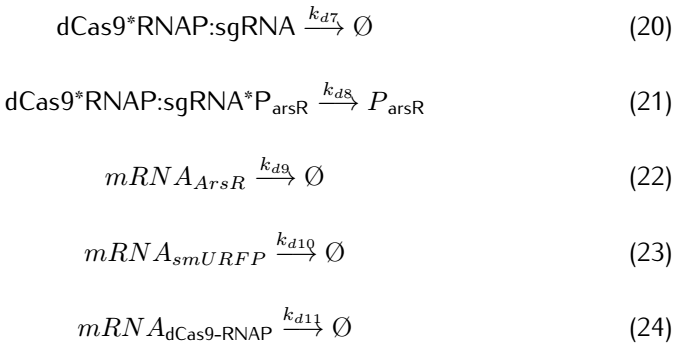
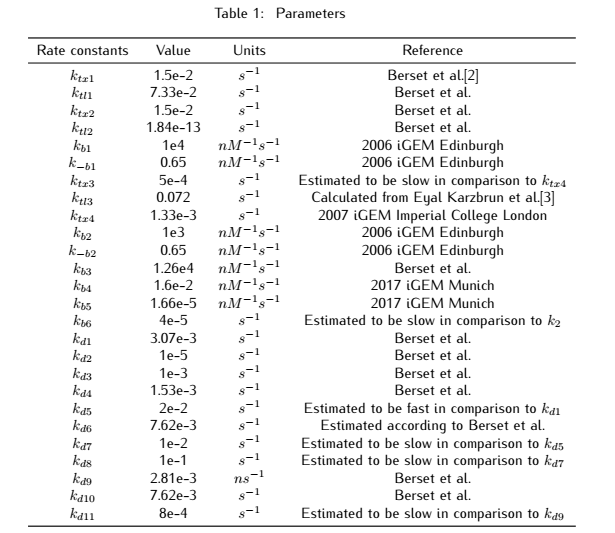
2.2 Analysis of ODEs
Applying mass action kinetic laws,we obtain the following set of differentiak equations.The several complexes involved:Ars$R^*$$P_{arsR}$,$As^{3+}$,${dCas9}^*$RNAP,${dCas9}^*$RNAP:sgRNA,${dCas9}^*$RNAP:${sgRNA}^*P_{arsR}$,
are respectively abbreviated as $cplx_1$,$cplx_2$,$cplx_3$,$cplx_4$,$cplx_5$.
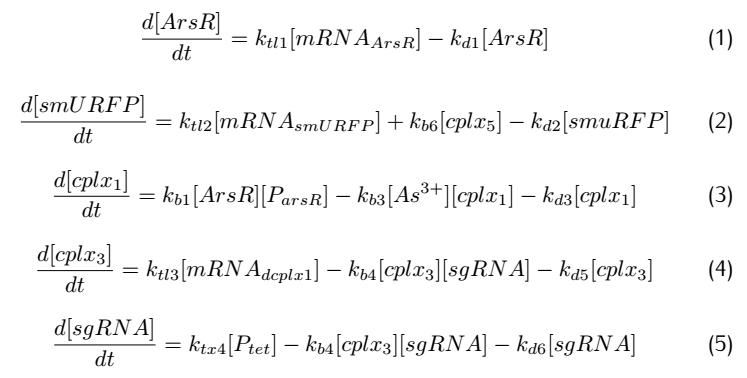
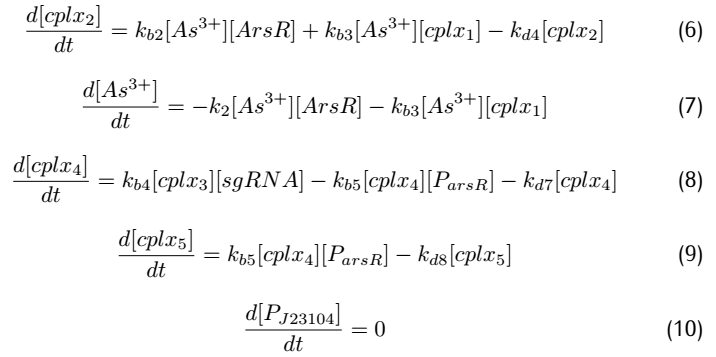

2.3 Simulation
Our simulation is based on two softwares: MATLAB (SimBiology Toolbox) and COPASI.
SimBiology Toolbox provides functions for modeling,simulating and analyzing biochemical pathways by the powerful computing engine of MATLAB.
SimBiology Toolbox provides functions for modeling,simulating and analyzing biochemical pathways by the powerful computing engine of MATLAB.
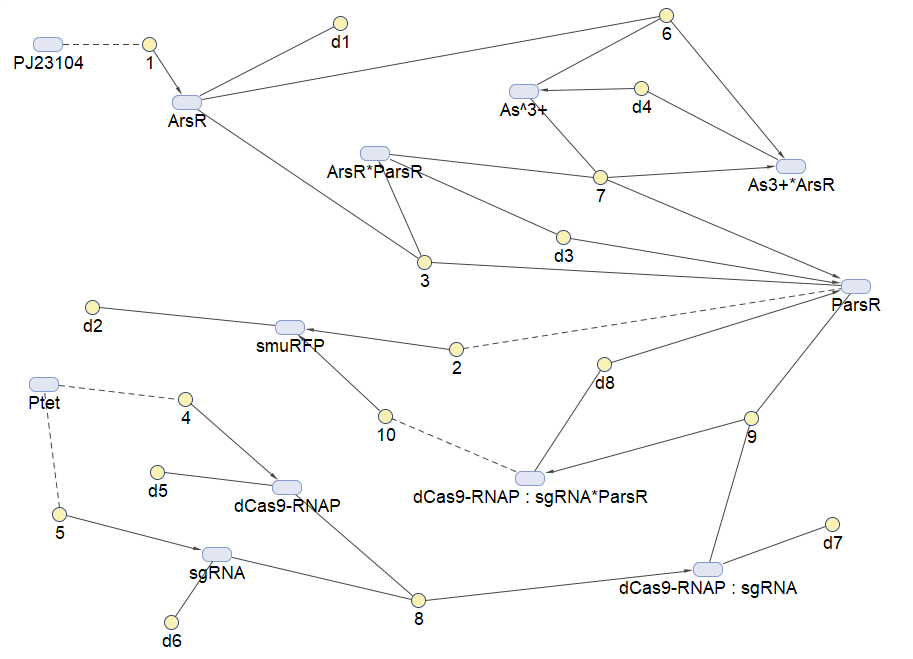
Figure 3:Reaction map generated from the reaction sets above by SimBiology Toolbox
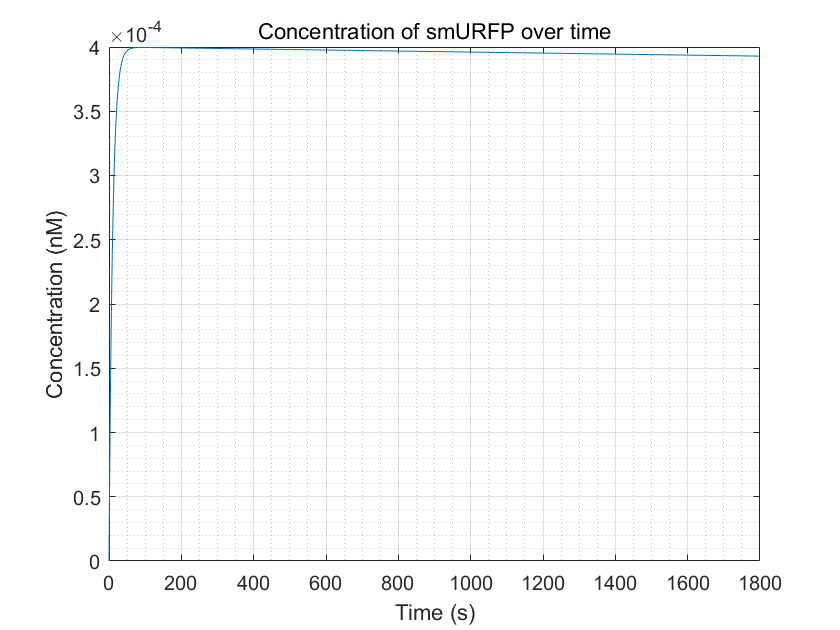
Figure 4:Simulation of smURFP production as a function of time by MATLAB Through the figure, we can see that the smURFP
can gradually increase and reach a steady state after a period in the presence of arsenic ions.
2.4 Sensitivity
A good biosystem should have certain stability towards fluctuations in parameters.A good model should reflect this,and
hence a test for robustness can be essential to the model.
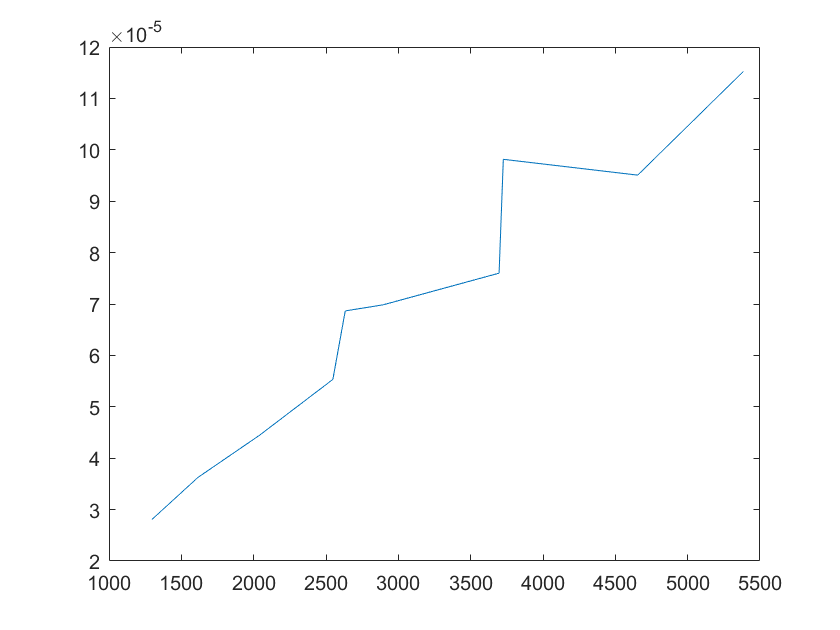
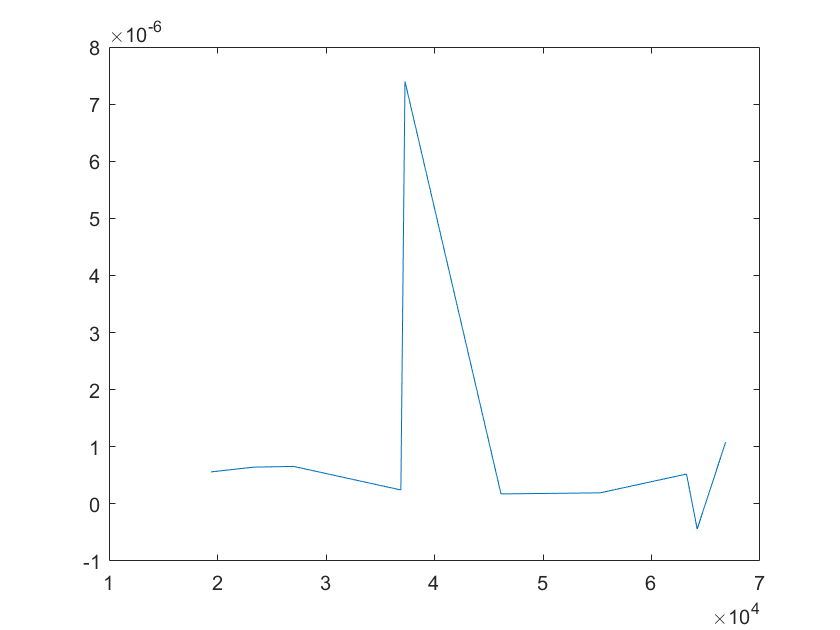
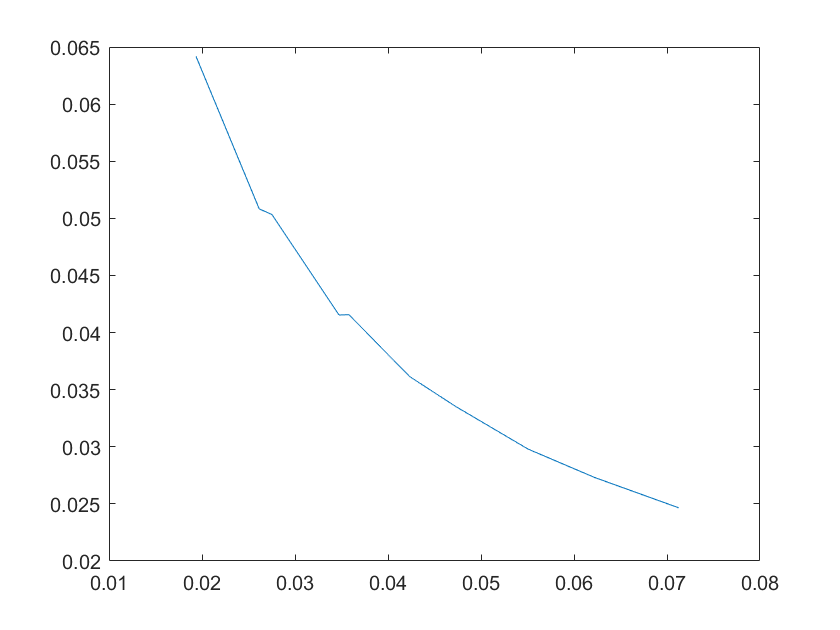
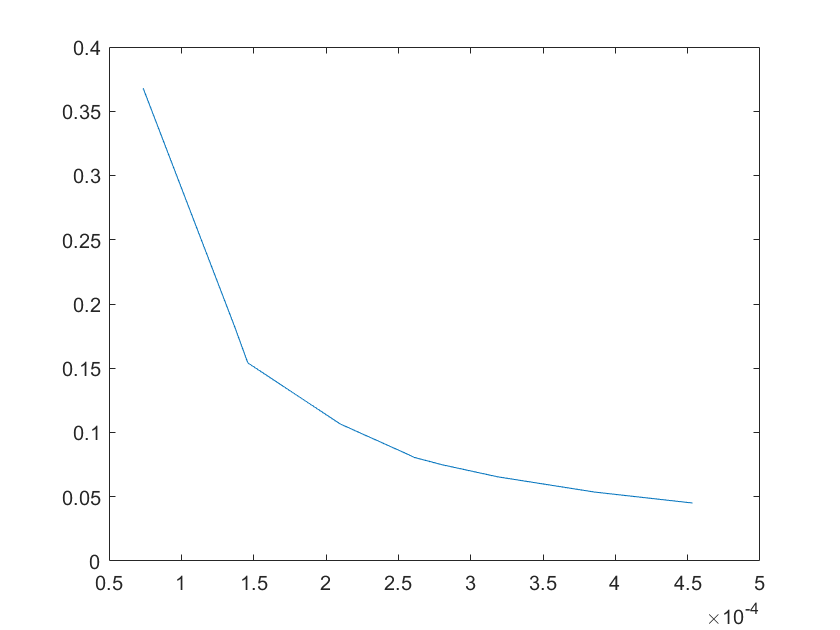
Robustness analysis can also pinpoint which reactions/parameters that are important for obtaining a specific biological behavior.A simple measure for sensitivity is to measure the relative change of a system feaure due to a change in a parameter.As for our model,the feature can be the equilibrium concentration of the smURFP(C) for which the sensitivity(S) to a parameter k is:
Robustness analysis can also pinpoint which reactions/parameters that are important for obtaining a specific biological behavior.A simple measure for sensitivity is to measure the relative change of a system feaure due to a change in a parameter.As for our model,the feature can be the equilibrium concentration of the smURFP(C) for which the sensitivity(S) to a parameter k is:
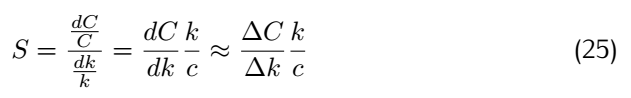
After analysis, we found that the concentration of smURFP is relatively sensitive to parameters such as ktx3,ktl3,ktx4,kb4,kb6,kd2,kd5,
kd6,kd7,kd8,kd11, etc. Among these parameters, except for the parameters that directly affect the production and
degradation of smURFP,the rest of them are all related to dCas9-RNAP:sgRNA. It shows that our model reflects the
critical role of dCas9-RNAP:sgRNA,which initially confirms our hypothesis:dCas0-RNAP can enhance transcription to
increase the concentration of smURFP. However, due to the lack of previous modeling studies on dCas9-RNAP,some kinetic
parameters may not be very accurate,and due to time limitation,we have not implemented experiments to measure related
parameters,which may lead to some deviations in our model.
The sensitivity of each parameter is shown in the figures below.
The sensitivity of each parameter is shown in the figures below.
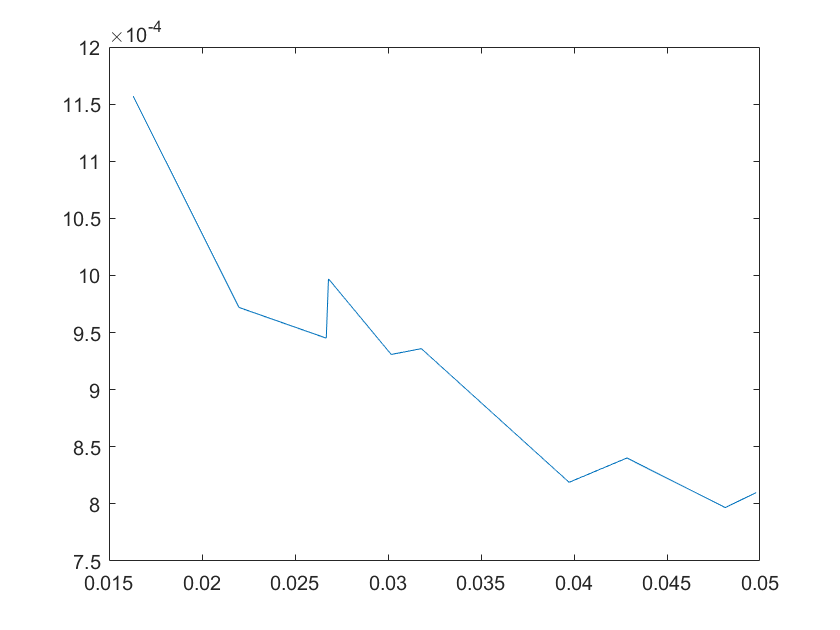
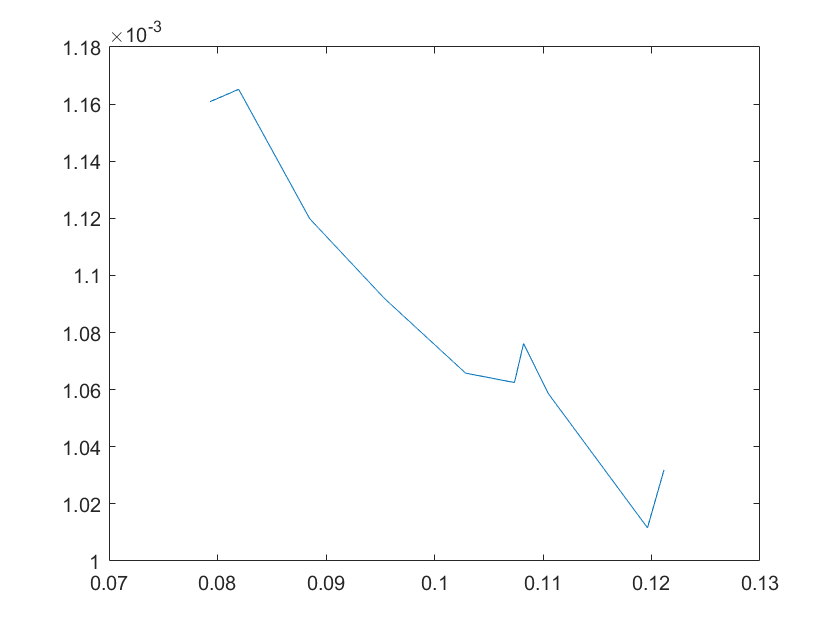
(a)sensitivity of ktx1 (b)sensitivity of ktl1
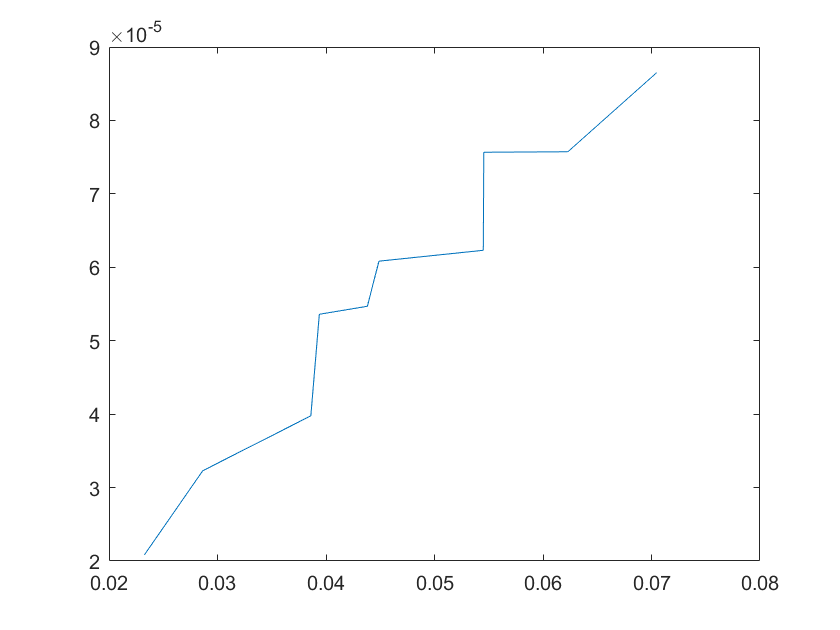
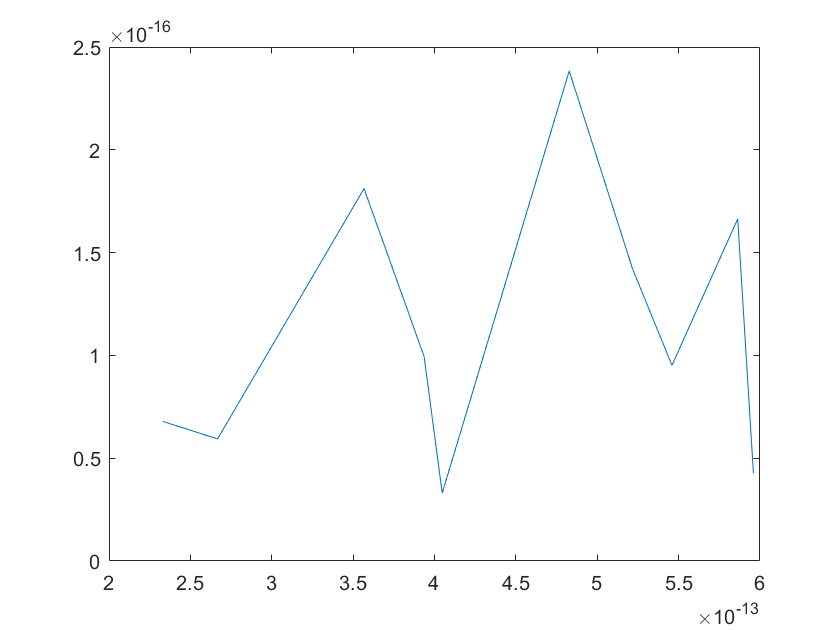
(c)sensitivity of ktx2 (d)sensitivity of ktl2
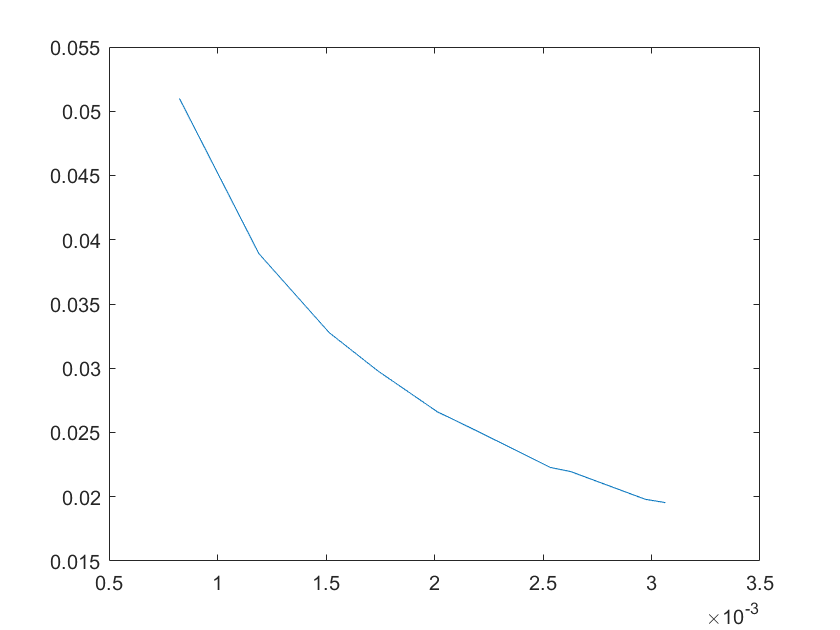
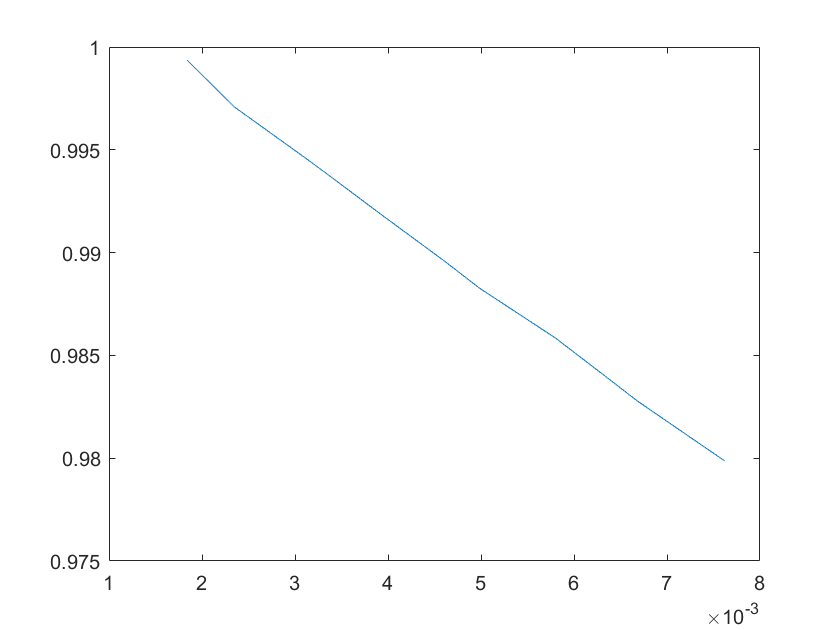
(e)sensitivity of ktx3 (f)sensitivity of ktl3
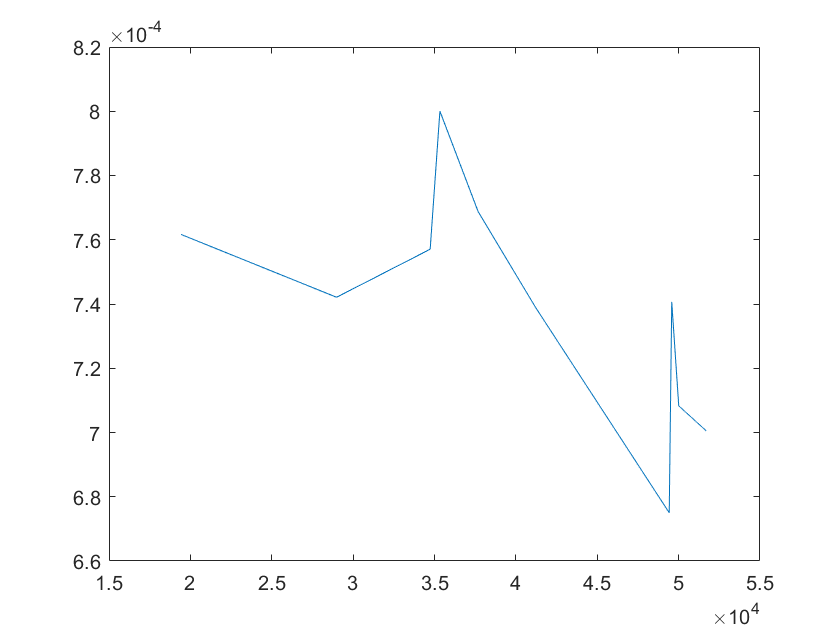
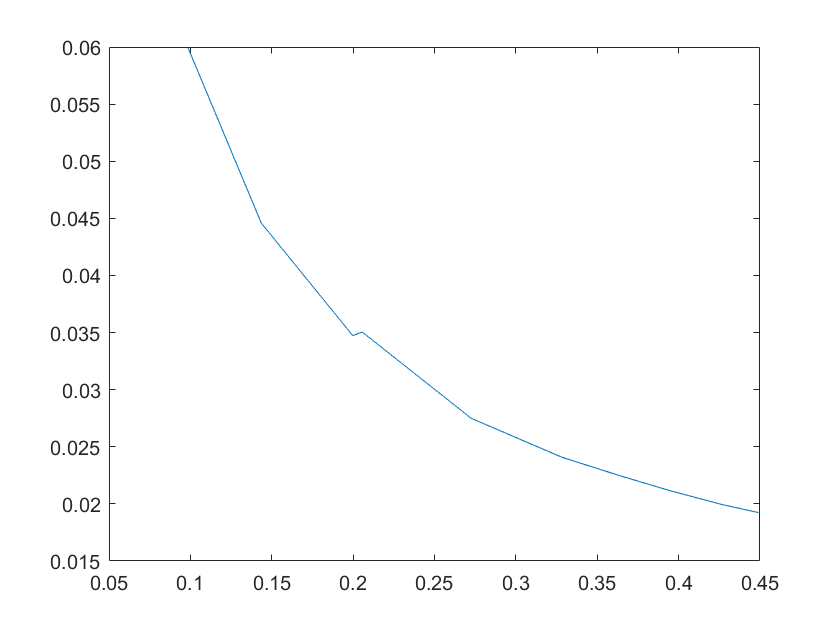
(g)sensitivity of ktx4 (h)sensitivity of kb1
(i)sensitivity of kb2 (j)sensitivity of kb3
(a)sensitivity of kb4 (b)sensitivity of kb5
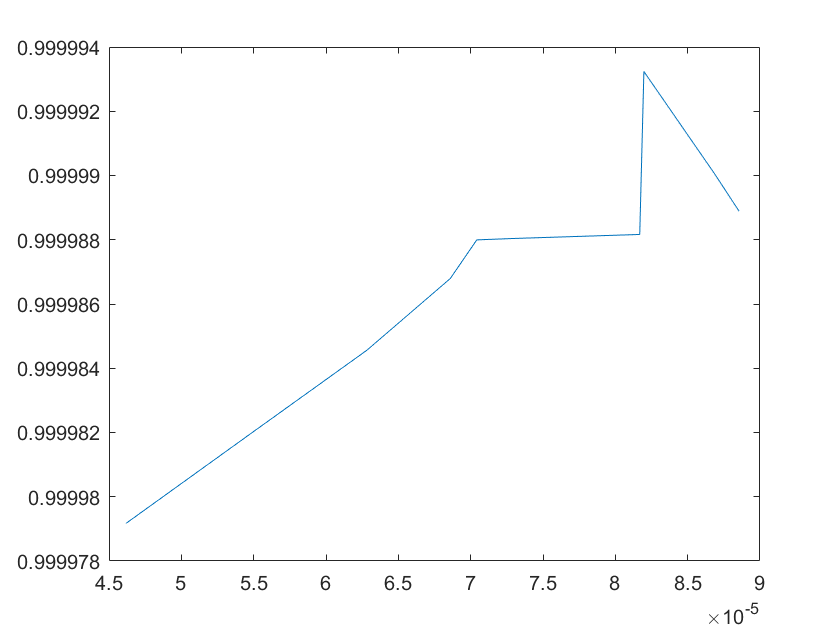
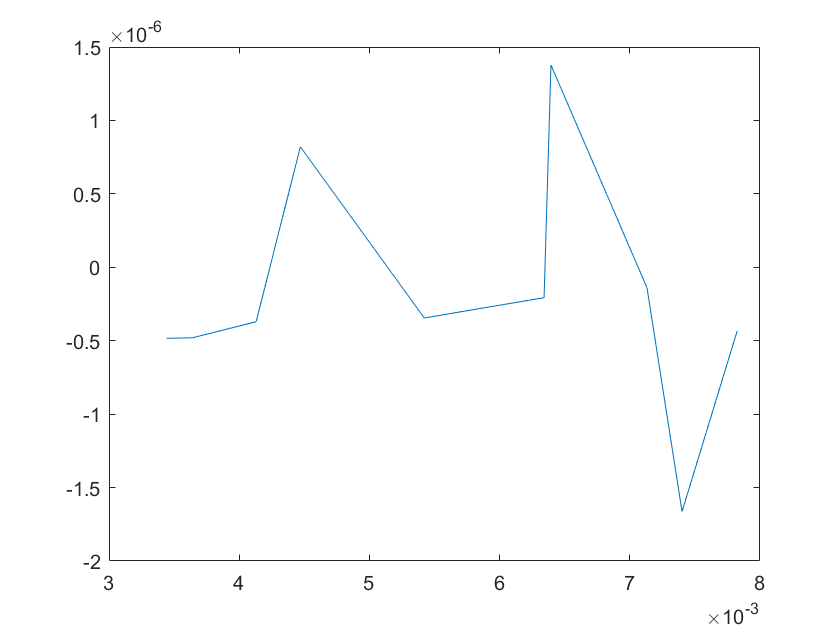
(c)sensitivity of kb6 (d)sensitivity of kd1
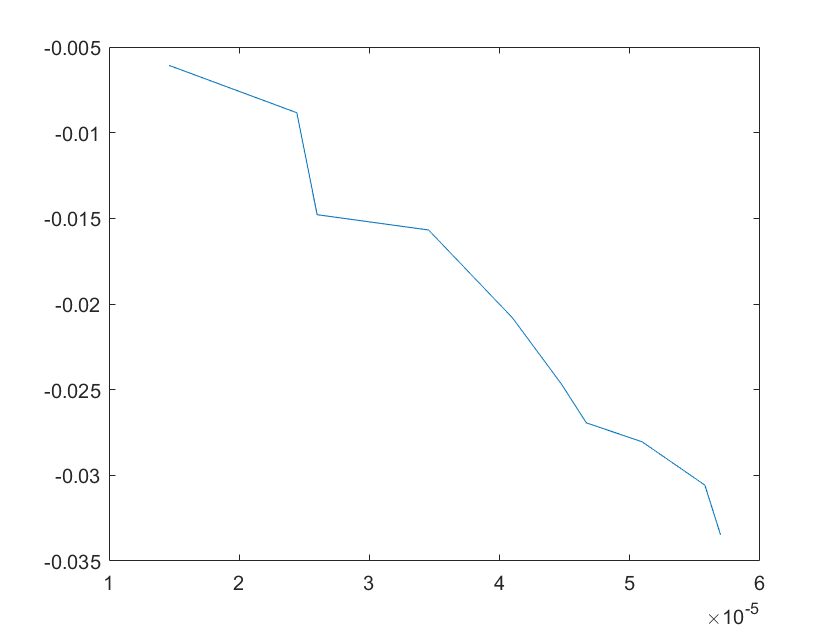
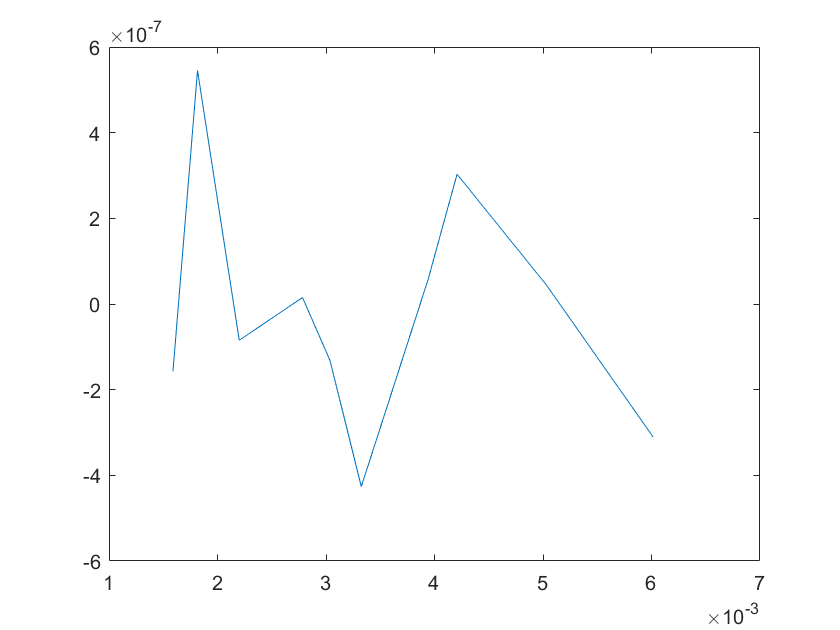
(e)sensitivity of kd2 (f)sensitivity of kd3
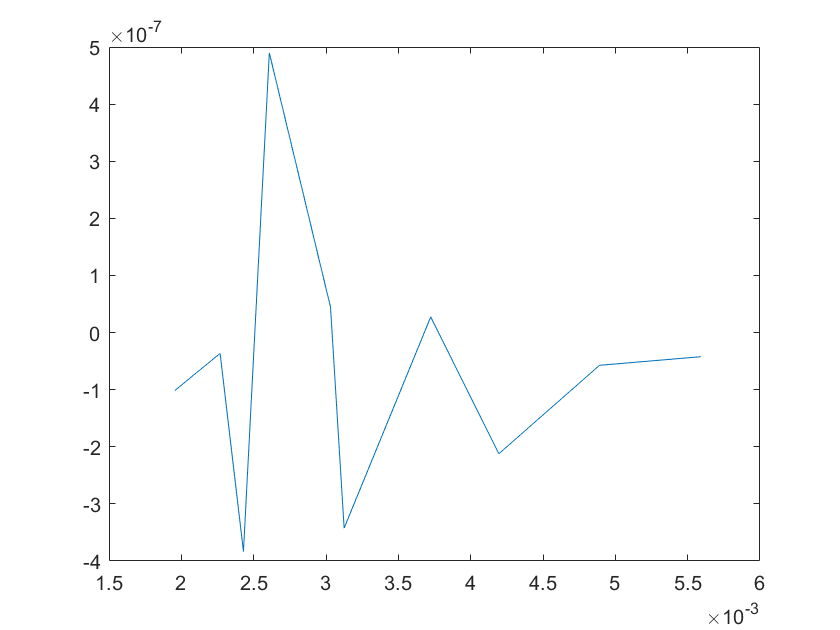
(g)sensitivity of kd4 (h)sensitivity of kd5
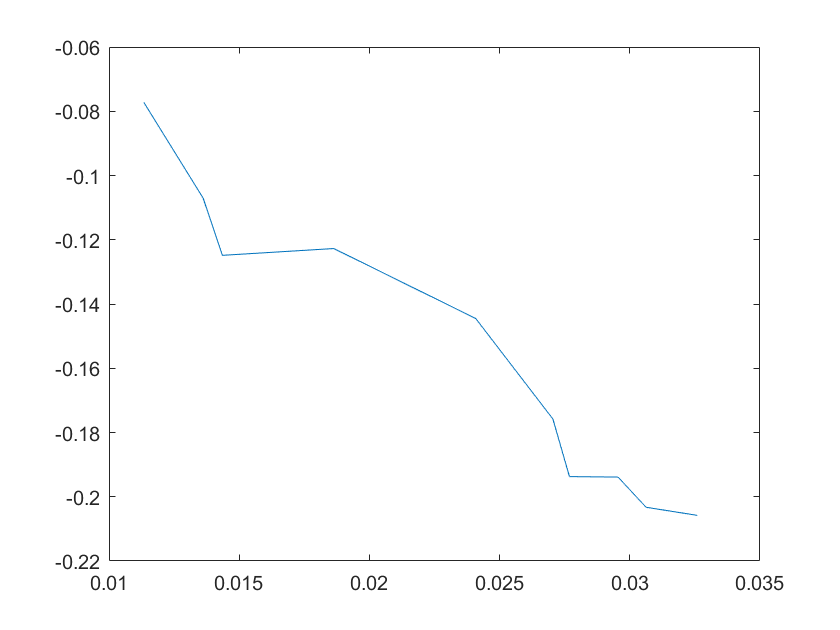
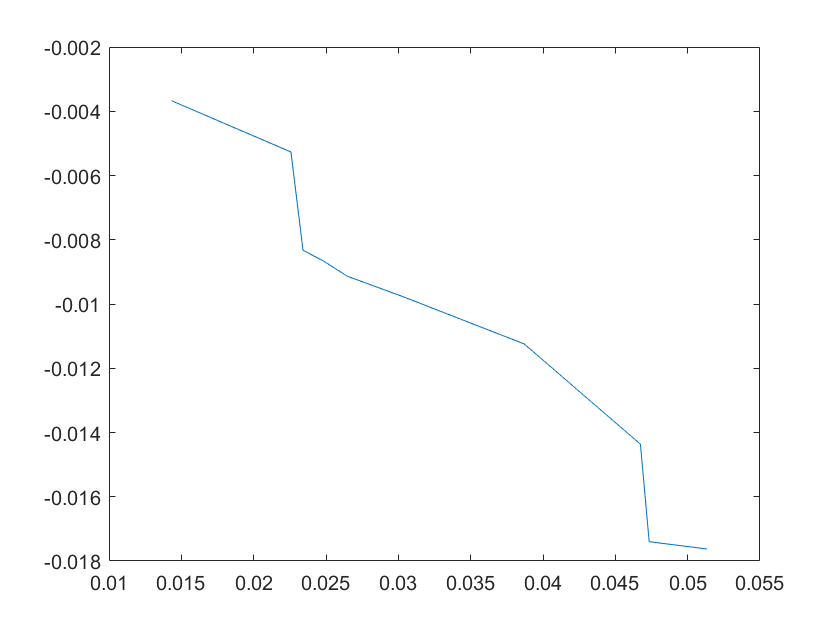
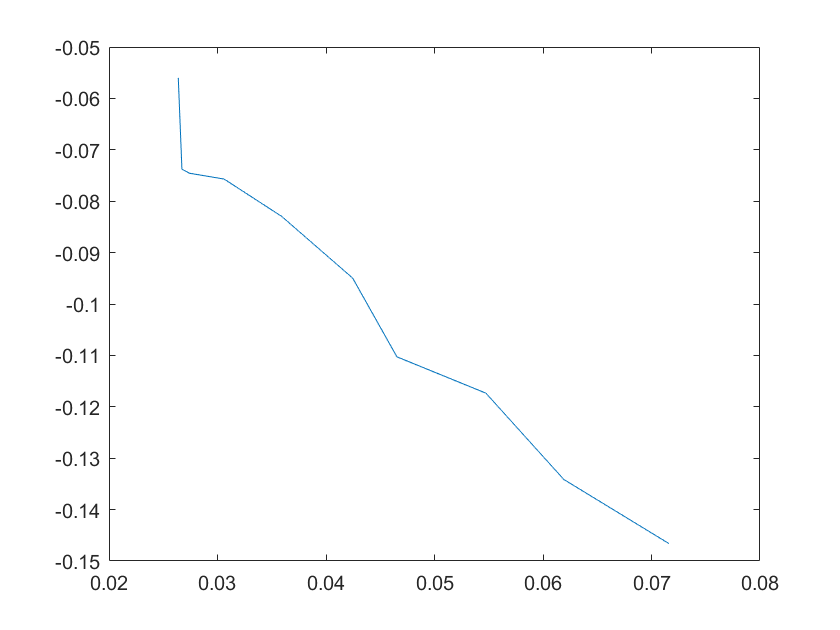
(a)sensitivity of kd6 (b)sensitivity of kd7
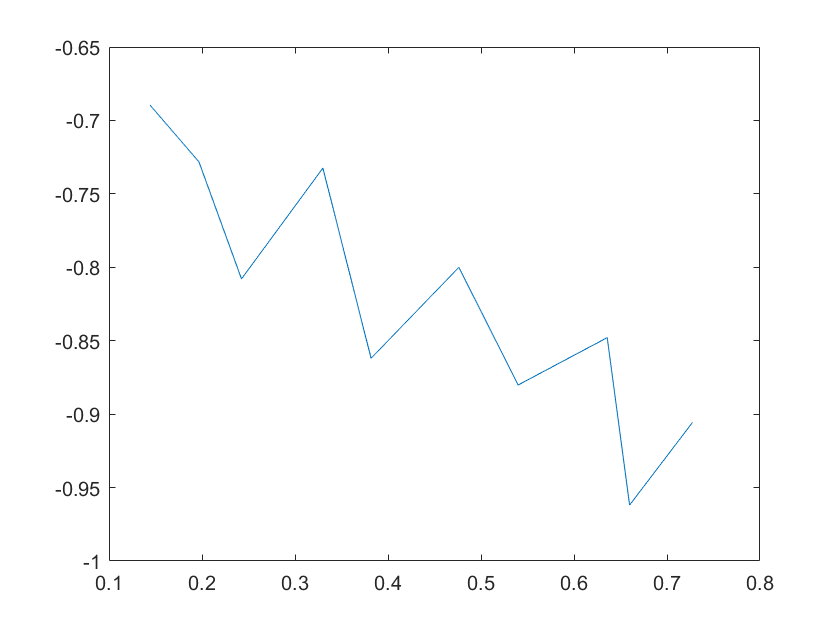
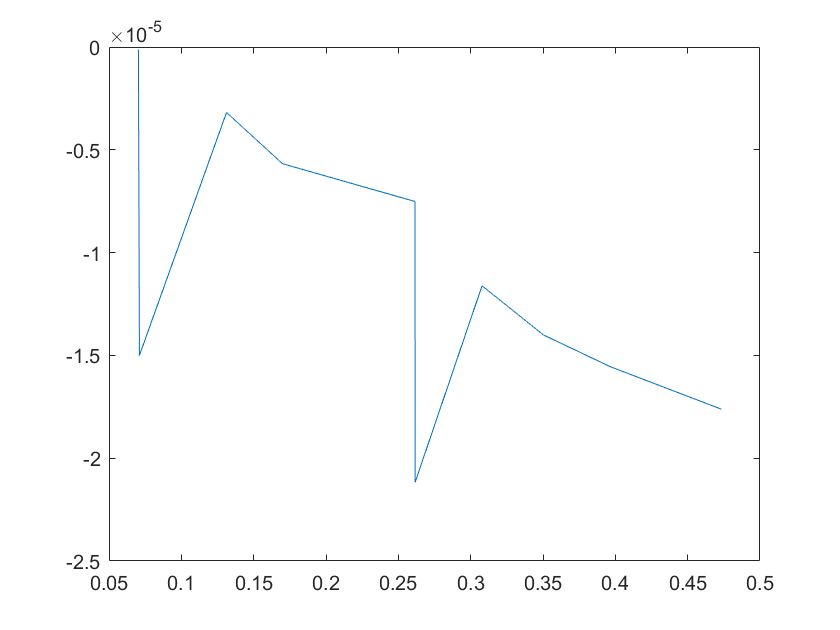
(c)sensitivity of kd8 (d)sensitivity of kd9
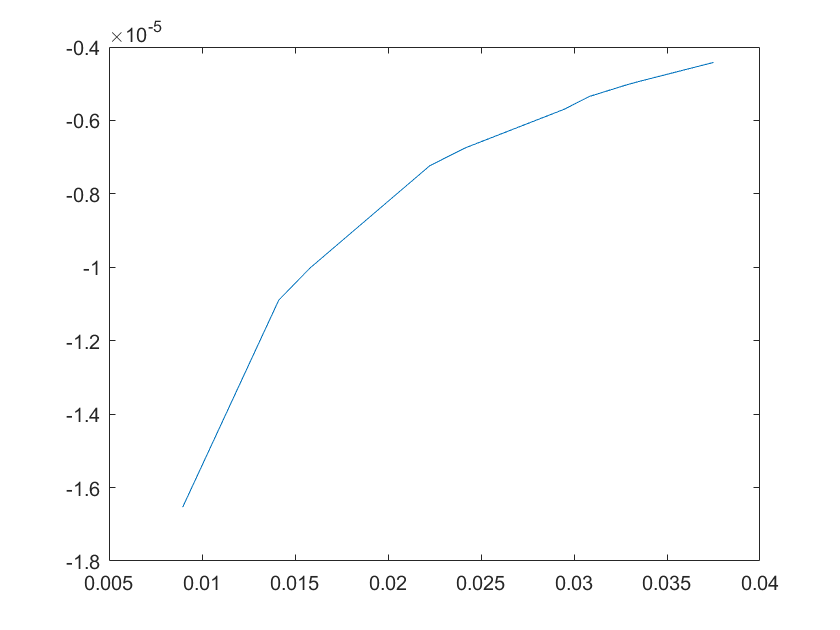
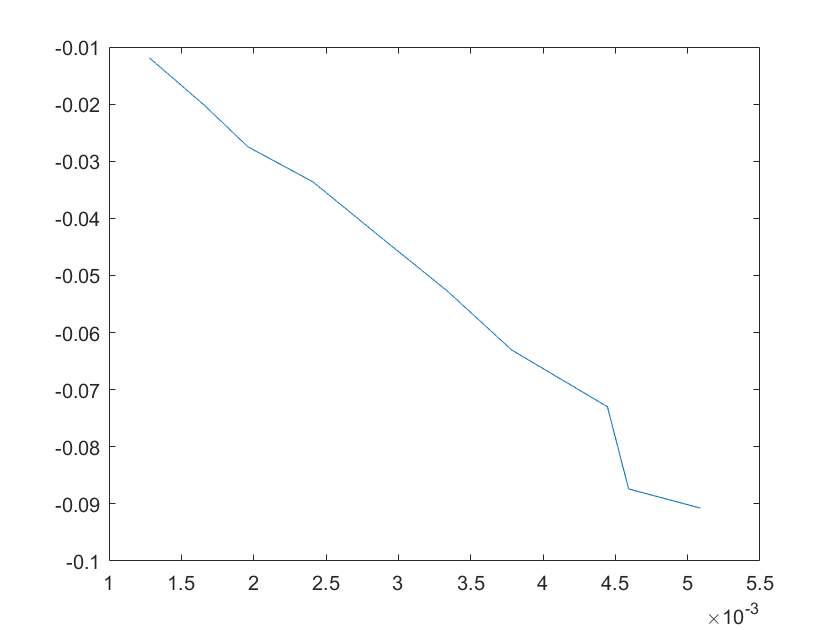
(e)sensitivity of kd10 (f)sensitivity of kd11
Note:The ordinate axis represents the sensitivity S,and the abscissa axis is the parameter k for which we want to evaluate
the sensitivity.
2.5 Application of the model
Since the goal of our project is to increase the sensitivity of biosensors by introducing a complex of dCas9-RNAP and
sgRNA, and one of the purposes of our model is to explore whether this complex is effective.So we assume a reasonable
and large enough concentration value for this complex. We use the concentration of Glyceraldehyde-3-phosphate dehydrogenase
A as the assumed concentration.Glyceraldehyde-3-phosphate dehydrogenase A(gapA) is a crucial enzyme in the glycolytic
pathway,and the gene encoding this enzyme is a housekeeping gene in E.coli cells with high expression levels.We find
in the literature that the protein mass of gapA is 48645 fg/cell,and its molecular weight is 35492 Da.[4] The amount
of abundance of Glyceraldehyde-3-phosphate dehydrogenase A protein per cell can be calculated as follows:
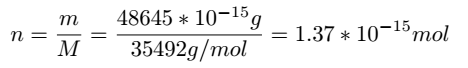
As for the size of E.coli,we found relevant data from the literature,as the figure below shows.[5]
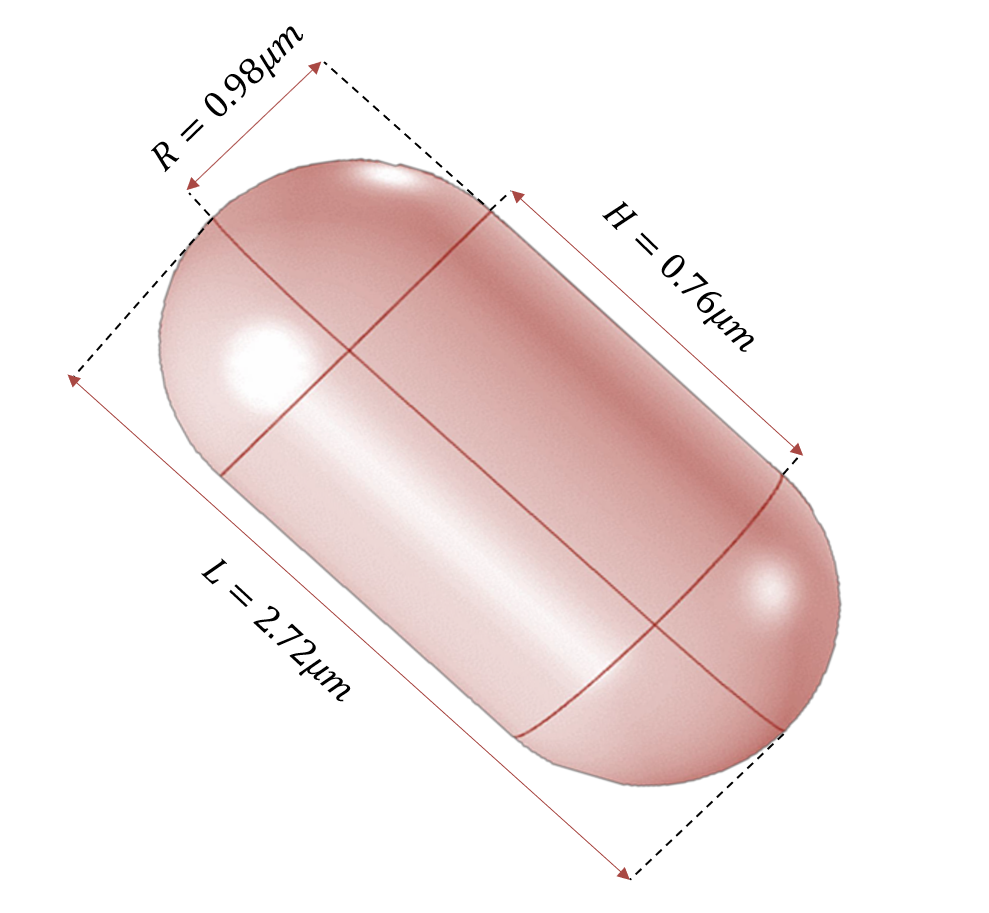
Figure 8:Size of E.coli
The volume of E.coli can be calculated as follows:
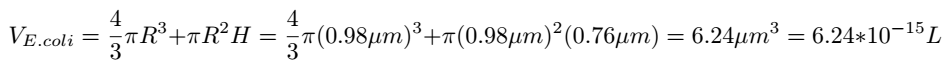
Then the concentration of Glyceraldehyde-3-phosphate dehydrogenase A protein in the cell can be determined:
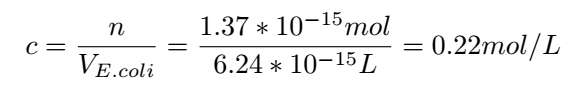
With this concentration,we can get very nice results:
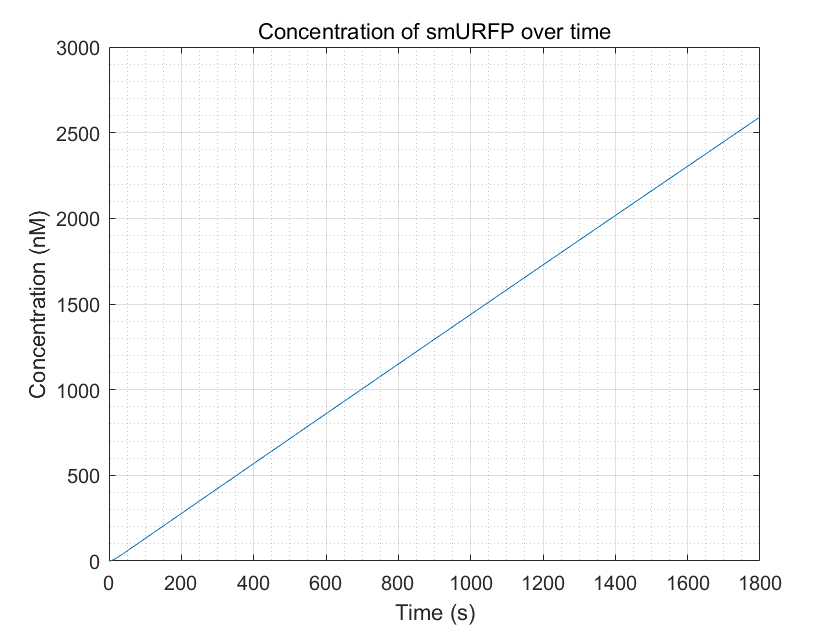
Figure 9:smURFP production with enough dCas9-RNAP:sgRNA
Compared to the diagram without introducing dCas9-RNAP:sgRNA:
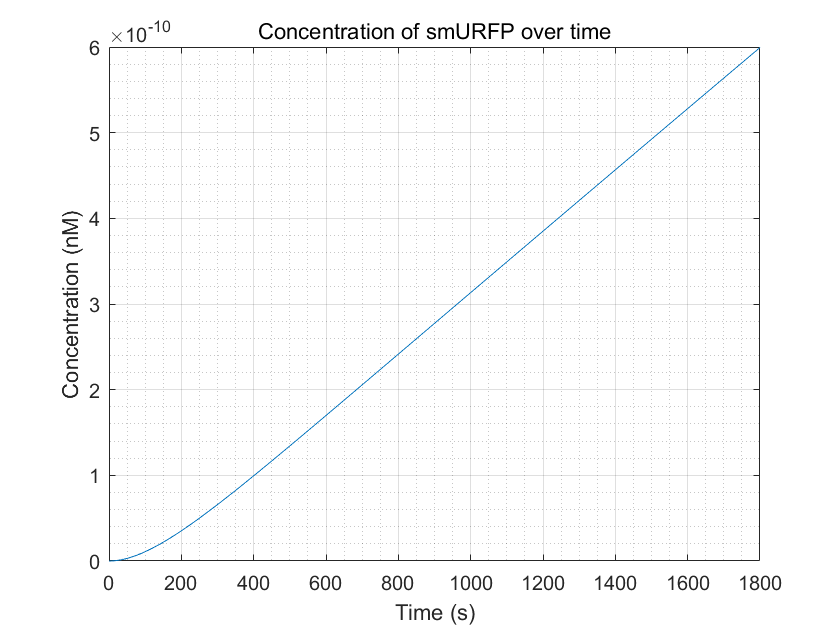
Figure 10:smURFP production within a reasonable time frame
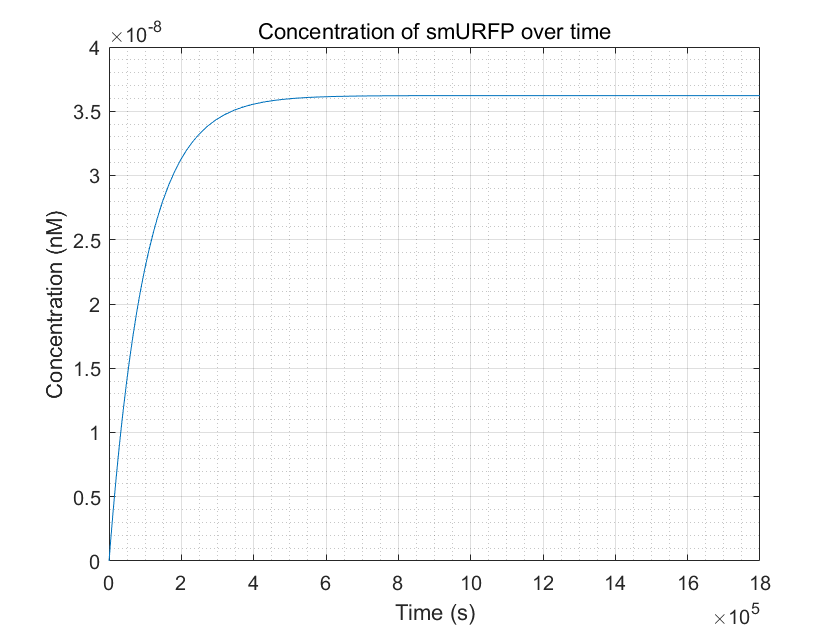
Figure 11:smURFP production reached equilibrium but it takes a long time
From these three figures, we can conclude that dCas9-RNAP:sgRNA does have the effect of promoting transcription and increasing
fluorescence intensity,thereby increasing sensitivity,as long as its concentration is sufficient.This result enhances
the confidence of the experimental group,and they need to try to improve the expression of dCas9-RNAP:sgRNA in E.coli
without having to doubt its role.
References
[1] LA Pola-Lopez et al."Novel arsenic biosensor "POLA" obtained by a genetically modified E.coli bioreporter cell" .In:Sensors
and Actuators B:Chemical254(2018),pp.1061-1068.
[2] Yves Berset et al."Mechanistic Modeling of Genetic Circults for ArsR Arsenic Regulation".In:ACS synthetic biology 6.5(2017),pp.862-874.
[3] Eyal Karzbrun et al."Coares-grained dynamics of protein synthesis in a cell-free system".In:Phtsical review letters 106.4(2011),p.048104.
[4] Yasushi Ishihama et al."Exponentially modified protein abundance index(emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein".In:Molecular E Cellular Proteomics 4.9(2005),pp.1265-1272.
[5] Nili Crossman,Eliora Z Ron,and Conrad L Woldringh."Changes in cell dimensions during amino acid starvation of Escherichia coli."In:Journal of bacteriology 152.1(1982),pp.35-41.
[2] Yves Berset et al."Mechanistic Modeling of Genetic Circults for ArsR Arsenic Regulation".In:ACS synthetic biology 6.5(2017),pp.862-874.
[3] Eyal Karzbrun et al."Coares-grained dynamics of protein synthesis in a cell-free system".In:Phtsical review letters 106.4(2011),p.048104.
[4] Yasushi Ishihama et al."Exponentially modified protein abundance index(emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein".In:Molecular E Cellular Proteomics 4.9(2005),pp.1265-1272.
[5] Nili Crossman,Eliora Z Ron,and Conrad L Woldringh."Changes in cell dimensions during amino acid starvation of Escherichia coli."In:Journal of bacteriology 152.1(1982),pp.35-41.
Construction of Free Energy Model
Zheng Hu,Sherry Dongqi Bao
TianJin University
October 10,2018
TianJin University
October 10,2018
1 Introduction
Nowadays,the analysis of cleavage possibility can be devided into two type,i,e.meta-empirical and empirical.For the first
one, people develop the various score function based on experiment data to evaluate if a sgDNA is good or bad.Correspondingly,the
other group chooce set up a theoretical model based on kinetic theory.But because using many approximations,it has
drawbacks inevitably.
Our model aims to investigate the off-target problem in gene editing by the CRISPR-Cas system,therefore finding efficient ways to enhance the reliability of gene editing.The foundations of thsi model are mostly simple probability theory and dynamic deduction,which make our model both convincing and pellucid.
Currently,people have constructed a similar model as illustrated in the following figure1.There are four common rules when Cas nuclease cleaves the DNA[1].
Our model aims to investigate the off-target problem in gene editing by the CRISPR-Cas system,therefore finding efficient ways to enhance the reliability of gene editing.The foundations of thsi model are mostly simple probability theory and dynamic deduction,which make our model both convincing and pellucid.
Currently,people have constructed a similar model as illustrated in the following figure1.There are four common rules when Cas nuclease cleaves the DNA[1].
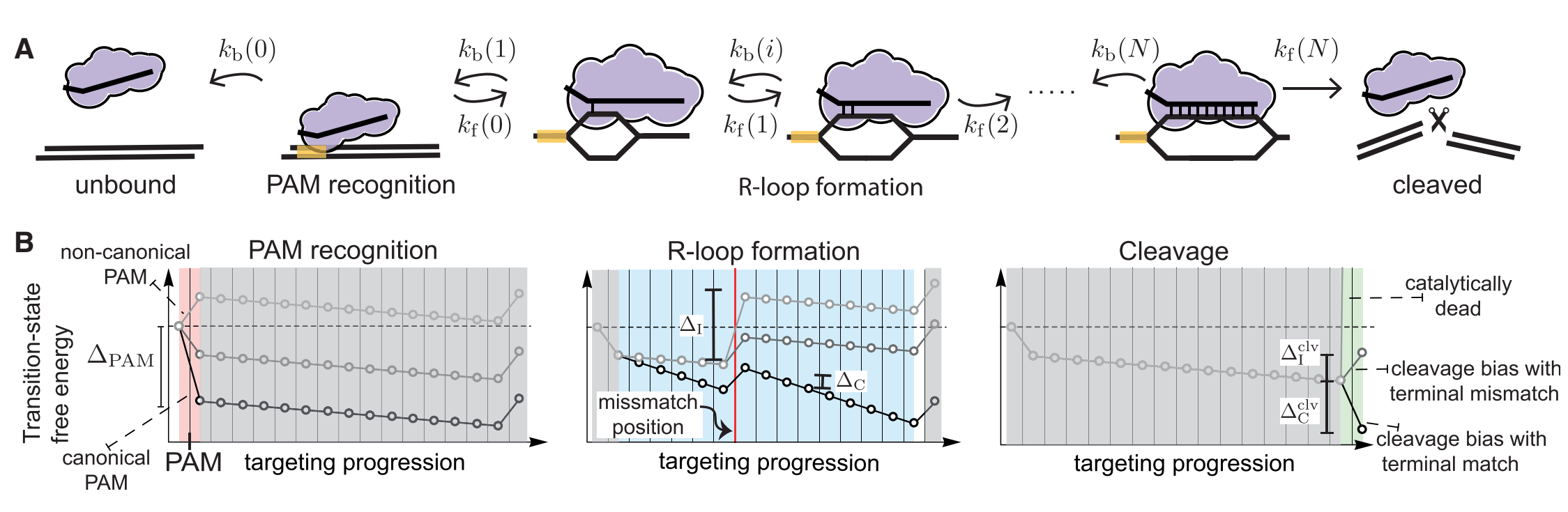
Figure q:schematic diagram
(1)Seed region:single mismatch(es) within a PAM proximal seed region can completely disrupt interference.
(2)Mismatch spread:when mismatches are outside the seed region,off-targets with spread out mismatches are targeted most strongly.
(3)Differential binding versus differential cleavage:binding is more tolerant of mismatched than cleavage.
(4)Specificity-efficiency decoupling:weakened protein-DNA interatctions can improve target selectivity while still maintaining efficiency.
Based on these four rules,probability theory is applied in to explain it.As we know,there are always only two results in an experiment,which are successful cleavage and unsuccessful cleavage.In math view,it can be one-hot encoded,and they are corresponding to 1 and 0.
(2)Mismatch spread:when mismatches are outside the seed region,off-targets with spread out mismatches are targeted most strongly.
(3)Differential binding versus differential cleavage:binding is more tolerant of mismatched than cleavage.
(4)Specificity-efficiency decoupling:weakened protein-DNA interatctions can improve target selectivity while still maintaining efficiency.
Based on these four rules,probability theory is applied in to explain it.As we know,there are always only two results in an experiment,which are successful cleavage and unsuccessful cleavage.In math view,it can be one-hot encoded,and they are corresponding to 1 and 0.

Figure 2

Figure 3
However,giving a 0/1 prediction is hard and unreliable.To solve this problem, one choice is to consider it as a cluster
problem;however,it is easier to find a continuous quantitative function rather than to find a suitable cluster distance
function.Sonaturally,finding an approximate probability distribution is a good choice.
In many target design toolkits,they use a score function with several param eters which can generate a score to evaluate whether the target is good or bad. Here we consider the score function has the similar ability to probability,which is a description of ”better” or ”worse” while can’t affirm whether successful cleavage willappear.For our case,our goal is to find a function indicating which target is BETTER.
Considering the difference between model prediction and experimental data,our model consists of two aspects,which are kinetic inference and an updating module.
In many target design toolkits,they use a score function with several param eters which can generate a score to evaluate whether the target is good or bad. Here we consider the score function has the similar ability to probability,which is a description of ”better” or ”worse” while can’t affirm whether successful cleavage willappear.For our case,our goal is to find a function indicating which target is BETTER.
Considering the difference between model prediction and experimental data,our model consists of two aspects,which are kinetic inference and an updating module.
2 Methods
2.1 Knietic module
Figure 2 shows that the whole binding-cleavage process begins with the bind ing between PAM andprotein.Therefore,it corresponds
to rule1 mentioned before.And as the reaction proceeds,every step of it is reversible,and its irre versibility mainly
depends on the binding energy of two DNA bases. The boundary probability Pclv;N,representing the probability of matching
at the Nth position(the last position of sgRNA) of nucleotide base,is given by:
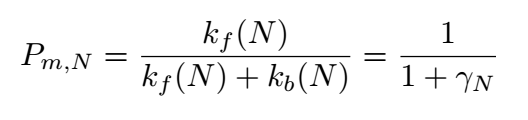

Figure 4

Figure 5
Where k is the reaction rate constant; f represents the forward reactions;b represents the backward reaction.And
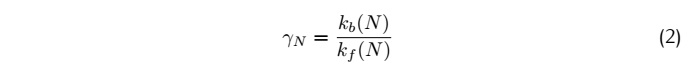
So for a complete match:
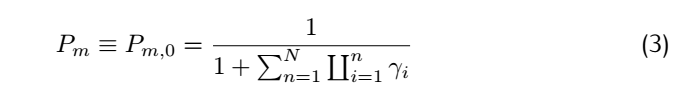
Consider the rate constant $K_f(i)$ and #k_b(i)$:
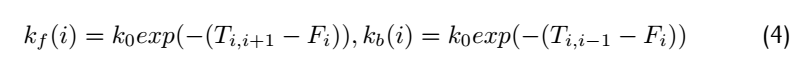
where $F_i$ means free energy of each metastable state,$T_{i,i+1}$means the highest free energy point on the reaction
path from position i to position i+1.Therefore,$T_{i,i+1}$-$F_i$ is the activation energy of forward reaction and
$T_{i,i+1}$-$F_i$ is activation energy of the backward reaction.
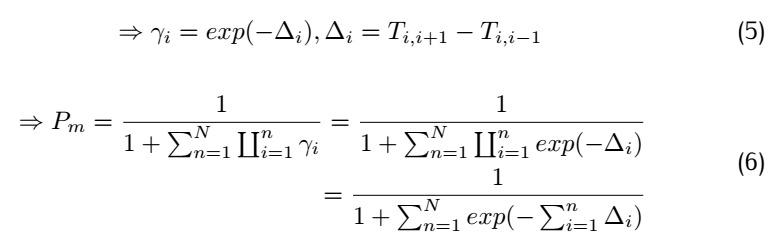
We define
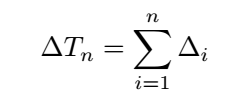
So
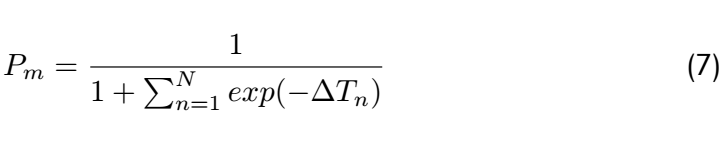
From the above,it is clear that the matching probability depends only on the state transition energy,not on the free
energy of the metastable states.If we assume there is one dominant minimal bias,say for n = n ∗ ,then this equation
can be approximated as:

To sum up,the cleavage possibility mainly relies on the free energy change, and PAM appears as a significant energy decline.
Figure 6:AT
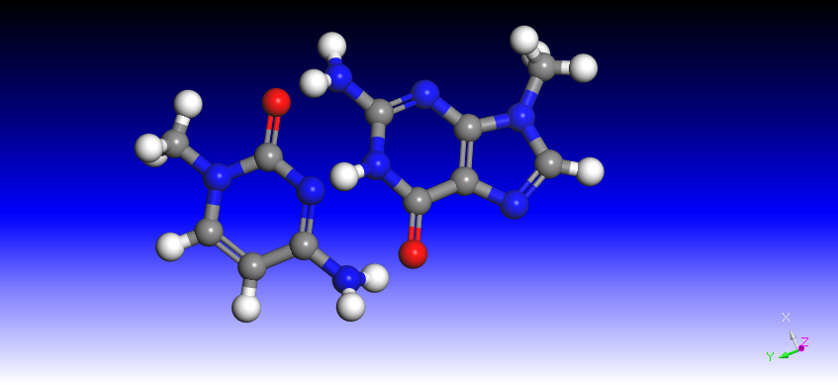
Figure 7:CG
So the kinetic module set up a form to regress the relationship between cleav age and the numbers of nucleotide matches
and mismatches.In consideration of this problem more carefully,the cleavage possibility becomes equal to analysis
energy change,and we know the binding energy of A/T and C/G is diferent due to the different hydrogen bond between
them.However,in appearing kinetic mod el,research tend to describe them in a rough definition as “matched base pairs”,
and the energy incline in C/G is approximately 1.5 folds as A/T.Similarly,the mismatch has more difference because
the size of nucleotides is various.Hence, the combination of the mismatched base pair was classified by group volume,i.e.
two pyrimidines(such as C/T,“L”),pyrimidine and purine(such as C/A,“M”),t wo purine(such as G/T,“S”).Hence,our possibility
can be calculated using the following formation.
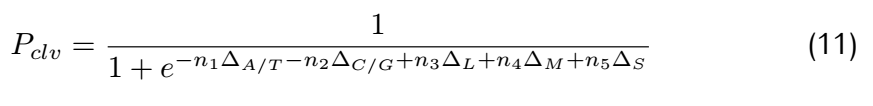
2.2 Optimization module
It is a common sense that experimental results are facts,but theoretical results are only conjectures.From kinetic module,we
can get an output,which is the cleavage possibility.The parameter we choose only aims to make results have discrimination,while
it’s not quantitative.And in a cleavage experiment,we only have two outcomes,successful and unsuccessful.To make
our prediction possibility more approximate toexperiment,we regard this as a regression problem.
Here,the method we choose is stochastic gradient descent(SGD) and cross entropy.And their principle can be concluded as follows.
Here,the method we choose is stochastic gradient descent(SGD) and cross entropy.And their principle can be concluded as follows.
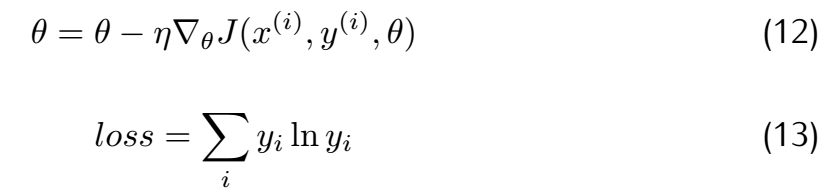
where θ means the parameters array and J means the loss function. Considering the difference in gradient calculation,we
use difference to substi tute differential aim to accelerate operating speed.
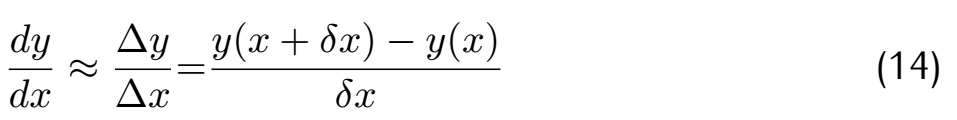
By using this simple method,our model can be more vibrant,updating using newest data and becoming more reliable.
2.3 Pre-selector
It’s obviously that the algorithm is too complex to applying in slide in a huge
DNA array.To solve this problem,we use a pre-selector to get some candidates
and use previous model to compare them so that we could get a greatest target.
And here this pre-selector structure is very simple.Considering use this map to reflect the similarity between target and full DNA.
And here this pre-selector structure is very simple.Considering use this map to reflect the similarity between target and full DNA.

Here,we use PAM as an input and collect the array which contain the same
beginning code as PAM.
3 Result
Here,we set the parameters as default values and observe its performance.As
the following figure shows,the energy always decreases or has some turning point
and is always negative.Such as the red line,it has a peak due to a mismatch
here,and in our model,we find that it doesn’t make the energy positive.That
means that in this reaction process there is some force like ”momentum” pushing
it to proceed and cross the energy peak.Corresponding to the other figure’s two
particular locations(aandb),only in these points their energy are all negative
(because we want to see the idea target series,only the locations which correspond
to negative energy are collected).After testing our code runtime,its manage rate
can reach approximate 2×$10^8$ base/h (under parallel calculation in 4 cores) and
have somewhat applicaiton value.Besides the default parameters,we hope our
model can hit more true data.So if we get the experiment data,we can use model
2.2 to get greater parameters.(@@no experiment data)
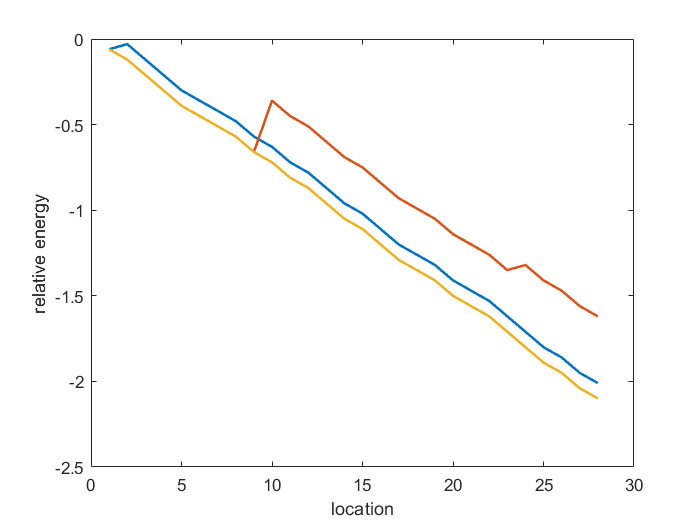
Figure 8 energy change
References
[1] family=Klein,familyi=K.,given=Misha,giveni=M.,"Hybridization kinet
ics explains CRISPR-Cas off-targeting rules".In:Cell reports 22.6(2018),pp.1413-1423.