Difference between revisions of "Team:Toulouse-INSA-UPS/Results"
| Line 163: | Line 163: | ||
<p> | <p> | ||
| − | We then incubated 3.2 µM of Orthos with 32 µM of <i>in vivo</i> biotinylated BFP. For the control experiment BFP (without Orthos), the same quantity of BFP was added to have a relevant comparison. These samples were then incubated with cellulose. After | + | We then incubated 3.2 µM of Orthos with 32 µM of <i>in vivo</i> biotinylated BFP. For the control experiment BFP (without Orthos), the same quantity of BFP was added to have a relevant comparison. These samples were then incubated with cellulose. After three washes with resuspension buffer, fluorescence was measured in the cellulose pellets. As shown in Figure 4, fluorescence is twice more intense in Orthos sample than in the control sample containing BFP alone. These experiments have been performed in quadruplicate and a Mann Whitney statistical test indicated that the increased fluorescence signal obtained with Ortos is significant. |
</p> | </p> | ||
| Line 183: | Line 183: | ||
<p> | <p> | ||
| − | Then, 58.0 µM of biotinylated FITC was incubated with 5.8 µM of Orthos protein. The same amount of FITC alone (58.0 µM) was used for the control experiment. These samples were incubated with cellulose and after | + | Then, 58.0 µM of biotinylated FITC was incubated with 5.8 µM of Orthos protein. The same amount of FITC alone (58.0 µM) was used for the control experiment. These samples were incubated with cellulose and after five washes with resuspension buffer, fluorescence levels were measured in the cellulose pellet fractions (Figure 5). In the presence of Orthos, fluorescence in the cellulose pellet was about twice higher than in control experiments corresponding to the cellulose pellet incubated with FITC alone (without Orthos) or with the reaction buffer only. These experiments was performed performed in quadruplicate and a Mann Whitney statistical test indicated that the increased fluorescence signal obtained with Ortos is significant. This result clearly shows that purified Orthos retains the ability to mediate the interaction between cellulose and a compound biotinylated <i>in vitro</i>. In addition, this result provides another strong evidence that Orthos allows to functionalize cellulose using a fluorescent molecule through its streptavidin linker. |
</p> | </p> | ||
| Line 275: | Line 275: | ||
</div> | </div> | ||
<p> | <p> | ||
| − | To test the potential of Cerberus in functionalizing cellulose, we monitored its ability to mediate an interaction between cellulose and a fluorescent compound. To generate a fluorescently labelled Cerberus protein, we performed a SPAAC reaction on 3.2 µM Cerberus protein using 31.9 µM µg of FITC-DBCO. In control experiment, 31.9 µM of FITC alone was used. These samples were then incubated with cellulose and after | + | To test the potential of Cerberus in functionalizing cellulose, we monitored its ability to mediate an interaction between cellulose and a fluorescent compound. To generate a fluorescently labelled Cerberus protein, we performed a SPAAC reaction on 3.2 µM Cerberus protein using 31.9 µM µg of FITC-DBCO. In control experiment, 31.9 µM of FITC alone was used. These samples were then incubated with cellulose and after five washes with resuspension buffer, fluorescence levels were measured in the cellulose pellet fractions (Figure 9). We observed that the fluorescence levels in the cellulose pellet incubated with Cerberus protein clicked to FITC were about 3 times higher than in the control experiments corresponding to the cellulose pellet incubated with FITC alone. These results show that Cerberus can efficiently be conjugated to fluorescent molecules bearing a DBCO group by click chemistry and that the resulting fluorescent molecules strongly interact with cellulose. Therefore, Cerberus is both a convenient and potent platform to functionalize cellulose. |
</p> | </p> | ||
| Line 281: | Line 281: | ||
<h5 id="Validation_Invitro_Cerberus" class="heavy">Validation using paramagnetic beads</h5> | <h5 id="Validation_Invitro_Cerberus" class="heavy">Validation using paramagnetic beads</h5> | ||
<p> | <p> | ||
| − | We further characterize the functionality of Cerberus in a second set of experiments using paramagnetic beads. To bind paramagnetic beads to Cerberus, we performed a click reaction using 3.2 µM of Cerberus protein and 32 µM DBCO-conjugated paramagnetic beads. As a control experiment, 32 µM of paramagnetic beads were used. These samples were then incubated with cellulose, and after | + | We further characterize the functionality of Cerberus in a second set of experiments using paramagnetic beads. To bind paramagnetic beads to Cerberus, we performed a click reaction using 3.2 µM of Cerberus protein and 32 µM DBCO-conjugated paramagnetic beads. As a control experiment, 32 µM of paramagnetic beads were used. These samples were then incubated with cellulose, and after five washes with resuspension buffer, we measured the magnetisation of cellulose using a magnet. |
</p> | </p> | ||
<div style="text-align:center"> | <div style="text-align:center"> | ||
Revision as of 21:23, 11 October 2018
RESULTS
INTRODUCTION

Sirius: CBM3a
Background
Sirius, named after the brightest star of the northern hemisphere alpha canis majoris, is a fusion protein between CBM3a and mRFP1. This design, which consists in fusing CBM3a to a fluorescent moiety, allowed us to investigate the binding capability of CBM3a platform to cellulose. This key step is a deliverable of our project, because only the CBM3a platform interacts with cellulose through four aromatic amino acids of its binding domain.
Key Achievements
-
Cloning of the part encoding Sirius
-
Overexpression and purification of Sirius
-
Specific fixation of Sirius to cellulose
Materials and Methods
The fused CBM3a and mRFP1 sequences were cloned into the pET28 expression vector in fusion with the His-tag. The resulting construct was transformed into E. coli strain and expression of the recombinant protein was induced using IPTG. The His-tagged protein was then purified on IMAC resin charged with cobalt and used in cellulose pull down assays. For the experimental details, see Experiments.
Results and Discussion
Production and purification of Sirius

As shown in Figure 1, we successfully purified Sirius protein. Indeed, induction with IPTG produced a large amount of a protein at the expected size for Sirius (52 kDa, lane CFE) that was then found predominant in elution samples (E1/40 and E1/100). We estimated the degree of purity of full length Sirius at about 72%. In addition to the full length protein, we observed several extra bands that likely correspond to proteolysis products.
Validation
Once produced in E. coli, fixation of Sirius to cellulose was tested using pull down assays. 70 µM of Sirius protein or mRFP1 (without CBM3a) or buffer were incubated with cellulose. After five washes with resuspension buffer, cellulose pellets were recovered and the associated fluorescence was measured.
Strikingly, only the cellulose pellet incubated with Sirius protein displays a high level of fluorescence (Figure 2). Control experiments showed that only background levels of fluorescence are retained in the cellulose pellet incubated with mRFP1 alone (no CBM3a). hese experiments have been performed in triplicate and statistical analyses indicate that the high level of fluorescence obtained with Sirius is significant. These results clearly show that CBM3a of Sirius interacts with cellulose, and thus mediates fixation of the mRFP1 protein to cellulose.

Orthos: Streptavidin and biotinylated compounds
Background
Once we proved the efficiency of the binding molecular platform of our system, CBM3a, we assessed the binding capacity of the first linker: the biotinylated molecule-binding module streptavidin.
Orthos, named after the guardian of Geryon's cattle, is a fusion protein between CBM3a and a monomeric streptavidin head. The strong affinity of streptavidin for biotin (dissociation constant of 10-13M) will allow to tightly bind biotinylated organic molecules to Orthos. We used two types of biotinylated compounds to monitor the ability of our platform to functionalize cellulose: fluorophores (mtag BFP and fluorescein) and antimicrobial peptide (scygonadin).
Key Achievements
-
Cloning of the part encoding Orthos
-
Production of Orthos carrying monomeric streptavidin
-
Production of biotinylated fluorophores in vitro and in vivo
-
Functionalization of cellulose with biotinylated compounds
Materials and Methods
The fusion between monomeric streptavidin and CBM3a was cloned into the pET28 expression vector. The resulting construct was transformed into E. coli strain BL21 and expression of the recombinant protein was induced using IPTG. The protein was then purified on Regenerated Amorphous Cellulose (RAC) and used in cellulose pull down assays. For the experimental details, see Experiments.
Results and Discussion
Purification of Orthos

Figure 3 presents the results of the purification of Orthos on RAC. The supernatant (S) contained all proteins produced in E. coli, and washes W1 and W2 allowed to eliminate most of non-specific interactions with cellulose. The elution step, using ethylene glycol, released Orthos protein from RAC. The band at about 40 kDa corresponds to Orthos (expected size: 39 kDa) and the band at 15 kDa likely corresponds to a proteolysis fragment of Orthos containing only the CBM3a module.
We estimated that the purification levels of the monomeric Orthos was about 45%, which is sufficient for our validation assays.
Validation of monomeric Orthos
Validation using in vivo biotinylation
In a first set of experiments, we assessed the ability of Orthos to functionalize cellulose with a compound biotinylated in vivo. For that purpose, we took advantage of a novel system for in vivo protein biotinylation that makes use of the biotin ligase BirA. Indeed, proteins containing the AviTag small peptide can be efficiently biotinylated in vivo by BirA. We thus co-expressed BirA and a version of the Blue Fluorescent protein (BFP) fused to the Avitag in E. coli. We added biotin in the culture medium during IPTG induction, in order to produce and purify biotinylated BFP.

We then incubated 3.2 µM of Orthos with 32 µM of in vivo biotinylated BFP. For the control experiment BFP (without Orthos), the same quantity of BFP was added to have a relevant comparison. These samples were then incubated with cellulose. After three washes with resuspension buffer, fluorescence was measured in the cellulose pellets. As shown in Figure 4, fluorescence is twice more intense in Orthos sample than in the control sample containing BFP alone. These experiments have been performed in quadruplicate and a Mann Whitney statistical test indicated that the increased fluorescence signal obtained with Ortos is significant.
These results demonstrate that Orthos protein binds efficiently to an in vivo biotinylated compound. In addition, they show that Orthos binds to cellulose and thus provide a proof of principle that Orthos design allows functionalization of cellulose with a fluorophore through its streptavidin linker.
Validation using in vitro chemical biotinylation
In a second set of experiments, we tested the ability of Orthos to functionalize cellulose with in vitro biotinylated compounds. We used the newly emerging technique Cu(I)-free strain-promoted alkyne-azide click chemistry (SPAAC) allowing to couple in vitro a molecule containing a dibenzocyclooctyne (DBCO) moiety to another molecule bearing an azide function. We used this technique to ligate in vitro biotin-DBCO to an azide-functionalized fluorescein (FITC), thus leading to the expected biotinylated FITC.

Then, 58.0 µM of biotinylated FITC was incubated with 5.8 µM of Orthos protein. The same amount of FITC alone (58.0 µM) was used for the control experiment. These samples were incubated with cellulose and after five washes with resuspension buffer, fluorescence levels were measured in the cellulose pellet fractions (Figure 5). In the presence of Orthos, fluorescence in the cellulose pellet was about twice higher than in control experiments corresponding to the cellulose pellet incubated with FITC alone (without Orthos) or with the reaction buffer only. These experiments was performed performed in quadruplicate and a Mann Whitney statistical test indicated that the increased fluorescence signal obtained with Ortos is significant. This result clearly shows that purified Orthos retains the ability to mediate the interaction between cellulose and a compound biotinylated in vitro. In addition, this result provides another strong evidence that Orthos allows to functionalize cellulose using a fluorescent molecule through its streptavidin linker.
Validation using scygonadin
In order to confirm that the streptavidin moiety of Orthos is able to interact with different kind of biotinylated compounds, we decided to bind it to an antimicrobial peptide, scygonadin. To biotinylate this compounds, we used the in vivo protein biotinylation system as the one used for BFP molecule. We co-expressed BirA with a version of scygonadin fused to the Avitag in Pichia pastoris.

A compléter!We then incubated Orthos with in vivo biotinylated scygonadin and we observed after incubation overnight halos of inhibition for each samples (Figure 6). We noticed an inhibition halo more important for the Orthos-scygonadin sample than the scygonadin alone control. Unfortunately, we repeated this test, but we did not obtained the same result. Therefore, we need to improve our experiment to prove the coupling between Orthos and biotinylated scygonadin. These results are nevertheless encouraging in the perspective of functionalizing cellulose using Orthos bound to antibiotic protein.
Perspectives
Our experiments using different kind of biotinylated compounds provide convincing proofs of concept of concept that Orthos can be conjugated, via its streptavidin head, to biotinylated organic molecules. The resulting molecules interact with cellulose and are therefore potent cellulose functionalizing compounds.
We can now envision to exploit the streptavidin linker of Orthos to ligate various compounds that can be biotinylated chemically (in vitro) or in vivo, via BirA co-expression.
In addition, we incorporated a TEV protease cleavage site between the N-terminus linker and the streptavidin. This allow to produce Orthos-bound biotinylated compounds that can be released by TEV cleavage if required, as in the case of odorant molecules for examples to a commercial use.
Cerberus: AzF
Background
Cerberus, named after the guardian of the gates of the Underworld, is our three-headed protein made of the cellulose-binding molecular platform (CBM3a) fused at the N-terminus to the biotinylated molecule-binding head (monomeric streptavidin) and at the C-terminus to the unnatural amino acid azido-L-phenylalanine (AzF). The incorporation of AzF is achieved through the recognition of the amber stop codon at the C-terminus of CBM3a by an orthogonal AzF-charged tRNA.
Coupling of molecules to the AzF head can be performed by click chemistry, either SPAAC or Cu(I)-catalyzed azide-alkyne click chemistry (CuAAC). To prove the versatility of the system, different compounds such as fluorescent proteins or paramagnetic beads have been clicked on Cerberus.
Key Achievements
-
Cloning of the part encoding Cerberus
-
Production of Cerberus
-
Functionalization of cellulose
Materials and Methods
The fusion between the streptavidin linker and CBM3a platform sequences followed by the amber stop codon was cloned into the pET28 expression vector in fusion with the His-tag. The resulting construct was co-transformed into E. coli with pEVOL expression vector (coding for amino acyl tRNA synthetase (aarS)/tRNA orthogonal pair for in vivo AzF incorporation in E. coli under L-arabinose induction) into E. coli strain and expression of the recombinant protein was induced using IPTG and L-arabinose, with addition of AzF in the medium. The His-tagged protein was then purified on IMAC resin charged with cobalt and used in cellulose pull down assays. For the experimental details, see Experiments.
Results and Discussion
Production and purification of Cerberus
Cerberus production requires a deferred induction. We first induced the synthesis of aaRS/tRNA pair with L-arabinose, and added AzF at the same time. One hour later, we induced the production of Cerberus with IPTG.

Cerberus production and purification were analysed on SDS gel (Figure 7). Induction in the presence of AzF (Figure 7, lane I+AzF) led to the expression of two major bands at 37 and 39 kDa approximately compared to control conditions, namely the non-induced (lane NI) and induced without AzF (lane I-AzF). We hypothesized that the upper band would correspond to Cerberus (expected molecular weight of monomeric streptavidin: 41 kDa), and the lower bands to Orthos proteins (Cerberus without the AzF and the His-tag). Surprisingly, both bands were present in the elution fractions which is incoherent since Orthos lacks an His-tag and was supposed to be washed out during purification steps. To confirm our hypothesis, we analysed the same fractions by western blot using antibodies detecting the His-tag (Figure 8).

Anti-His-tag antibodies revealed a band at about 45 kDa in the sample corresponding to IPTG induction in the presence of AzF (Figure 8, lane I+AzF). This band is not present in control samples (Figure 8, lane NI and I-AzF), indicating that it corresponds to Cerberus protein (theoretical size: 41 kDa). In addition to the full length protein , we observed several extra bands which very likely correspond to proteolysis products since they are detected with the anti-His-tag antibodies. Moreover, the band at 45 kDa is clearly detected in elution samples (Figure 8, lanes E). We estimated that the purification level of Cerberus with monomeric streptavidin in the elution samples was about 62%. These data show that Cerberus was efficiently purified and can be used for pull down assays. In addition, these results show that the experimental setup to produce Cerberus also leads to the production of Orthos when the amber stop codon is not recognized by the AzF-charged orthogonal tRNA. Although Orthos does not contain a His-tag at its C-terminus, the protein seems to be efficiently co-purified with Cerberus. The basis of this observation is unclear but it may suggest that Orthos and Cerberus interact with each other, via their CBM3 or streptavidin moieties.
Validation
Validation using FITC (Fluorescein isothiocyanate) molecules
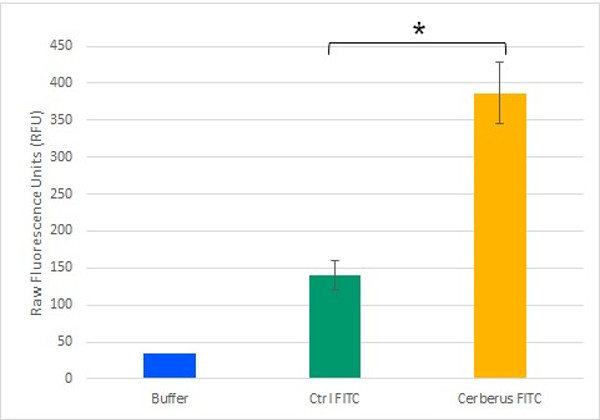
To test the potential of Cerberus in functionalizing cellulose, we monitored its ability to mediate an interaction between cellulose and a fluorescent compound. To generate a fluorescently labelled Cerberus protein, we performed a SPAAC reaction on 3.2 µM Cerberus protein using 31.9 µM µg of FITC-DBCO. In control experiment, 31.9 µM of FITC alone was used. These samples were then incubated with cellulose and after five washes with resuspension buffer, fluorescence levels were measured in the cellulose pellet fractions (Figure 9). We observed that the fluorescence levels in the cellulose pellet incubated with Cerberus protein clicked to FITC were about 3 times higher than in the control experiments corresponding to the cellulose pellet incubated with FITC alone. These results show that Cerberus can efficiently be conjugated to fluorescent molecules bearing a DBCO group by click chemistry and that the resulting fluorescent molecules strongly interact with cellulose. Therefore, Cerberus is both a convenient and potent platform to functionalize cellulose.
Validation using paramagnetic beads
We further characterize the functionality of Cerberus in a second set of experiments using paramagnetic beads. To bind paramagnetic beads to Cerberus, we performed a click reaction using 3.2 µM of Cerberus protein and 32 µM DBCO-conjugated paramagnetic beads. As a control experiment, 32 µM of paramagnetic beads were used. These samples were then incubated with cellulose, and after five washes with resuspension buffer, we measured the magnetisation of cellulose using a magnet.
As observed on the video, the cellulose incubated with the Cerberus protein conjugated to paramagnetic beads is quickly and totally attracted by the magnet. In contrast, in the control experiment, we only observed a slight movement of a part of the cellulose towards the magnet. This can be explained by the fact that washes do not remove all paramagnetic beads non-attached to the cellulose, or that paramagnetic beads naturally bind to cellulose but this interaction is not strong enough to resist washes.
These results show that Cerberus has the ability to interact simultaneously with cellulose and molecules with DBCO group, indicating that this modified protein, allows to functionalize cellulose through its linker containing unnatural amino acid (AzF).
Compounds functionalization
To go further in our project, we decided to functionalyse two inorganic molecules, graphene and carbon nanotubes, using a reaction of diazotization. This experiment was performed with the help of the ENSIACET laboratory in Toulouse. To check functionalization, we analysed samples by Thermogravimetric Analysis (TGA). The concept of TGA consists in measuring the mass change of a sample depending on temperature and time, in a controlled atmosphere. We choose to perform the experiment under azote, an inert gas that avoids mass loss by oxidation, and we studied the mass change from 25 to 1,000 °C (15 °C/min).


Figures 10 and 11 present the results of mass loss for functionalized Carbon Nanotube (CNT) and non-functionalized CNT samples, respectively. For functionalized CNT sample (Figure 10), we observed a mass loss of 27.4% around 100 °C corresponding to water loss. A second step of mass loss is observed around 550 °C, with a diminution of 19.8%. For the non-functionalized sample (Figure 11), we observed two mass losses at 150 and 750 °C corresponding to 1.5% and 3% of mass loss respectively. The significative difference between these two samples allowed us to conclude that the CNT functionalization has been successful.


Figures 12 and 13 present the results of mass loss for functionalized graphene and non-functionalized graphene samples respectively. For functionalized graphene sample (Figure 12), we observed a mass loss of 31.0% around 100 °C corresponding to water loss. A second step of mass loss is observed around 500 °C, with a diminution of 19.0%. For the non-functionalized sample (Figure 13), we did not observe a significative mass loss. So we can conclude that the graphene functionalization has been successful.
Perspectives
Our experiments provide robust proofs of concept that Cerberus can be conjugated, via its unnatural amino acid AzF, to organic or inorganic molecules bearing a DBCO group by click chemistry. The resulting molecules strongly interact with cellulose and are therefore potent cellulose functionalizing compounds.
We can now consider fixation of various compounds that can be chemically functionalized. We can also imagine double fixations, for example a fluorophore on the streptavidin head and paramagnetic beads on the AzF linker. This double fixation allows to envision endless possibilities to functionalize cellulose.
Bacterial Cellulose production and functionalization
Background
Some microorganisms naturally produce cellulose. One of these is Gluconacetobacter hansenii, a species of acetic acid bacteria, which can produce cellulose as a biofilm to protect itself. We chose to work with this bacterium to produce cellulose, because the iGEM Imperial team 2014 has set up culture conditions to optimize cellulose production. We used the best culture conditions to obtain a maximum of cellulose.
Key Achievements
-
Production of bacterial cellulose
-
Functionalization of bacterial cellulose with Sirius
Materials and Methods
G. hansenii was grown in Hestrin-Shramm (HS) medium at 30 °C. After one week of culture, bacterial cellulose was extracted and purified. For the experimental details, see Experiments.
For the functionalization, bacterial cellulose was incubated with purified Sirius or with purified mRFP1 alone as a control. Cellulose was then washed several times with resuspension buffer to remove non-attached proteins.
Results and Discussion

Figure 14 presents the results of the functionalization of our bacterial cellulose with a fluorescent protein. Only the cellulose sample incubated with Sirius (CBM3a-mRFP1) remains fluorescent after several washes with resuspension buffer. The control cellulose samples showed that only a weak coloration is retained after incubation with mRFP1 alone (not coupled with CBM3a platform). These results show that CBM3a interacts with the bacterial cellulose, and allows fixation of the mRFP1 protein to cellulose.
Perspectives
Now that we proved the capacity of CBM3a to interact with bacterial cellulose, we are confident that Orthos and Cerberus proteins should also interact with bacterial cellulose. Moreover, since we proved that we can conjugate several organic and inorganic compounds to the binding modules of Cerberus (streptavidin and AzF), we can envision functionalizing bacterial cellulose with many different compounds to provide it with new features.
No dogs were harmed over the course of this iGEM project.
The whole Toulouse INSA-UPS team wants to thank our sponsors, especially:









And many more. For futher information about our sponsors, please consult our Sponsors page.
The content provided on this website is the fruit of the work of the Toulouse INSA-UPS iGEM Team. As a deliverable for the iGEM Competition, it falls under the Creative Commons Attribution 4.0. Thus, all content on this wiki is available under the Creative Commons Attribution 4.0 license (or any later version). For futher information, please consult the official website of Creative Commons.
This website was designed with Bootstrap (4.1.3). Bootstrap is a front-end library of component for html, css and javascript. It relies on both Popper and jQuery. For further information, please consult the official website of Bootstrap.