Difference between revisions of "Team:SCAU-China/Demonstrate"
| Line 168: | Line 168: | ||
margin: 0; | margin: 0; | ||
| − | min-height: | + | min-height: 9000px; |
padding-top: 70px; | padding-top: 70px; | ||
min-width: 1140px; | min-width: 1140px; | ||
| Line 180: | Line 180: | ||
#mask { | #mask { | ||
width: 100%; | width: 100%; | ||
| − | height: | + | height: 9000px; |
background: rgba(0, 0, 0, 0.3); | background: rgba(0, 0, 0, 0.3); | ||
margin-top: -15px; | margin-top: -15px; | ||
| Line 443: | Line 443: | ||
<div class="DBoard" id="title" style="height:40px;width:400px;font-size:20px;line-height:40px;text-align:left;"> | <div class="DBoard" id="title" style="height:40px;width:400px;font-size:20px;line-height:40px;text-align:left;"> | ||
1.Genetic transformation of cyanobacteria</div> | 1.Genetic transformation of cyanobacteria</div> | ||
| − | <div class="DBoard" id="article1" style="height: | + | <div class="DBoard" id="article1" style="height:430px;"><!-- 展板 --> |
<p>Dry land growth cyanobacteria have strongly abilities to survive in desert. Therefore, it can be used as an ideal bioreactor in for producing cellulose which is a good material for water retention. We cloned and over-expressed seven bacterial genes (BCS genes) which are responsible for cellulose synthesis into cyanobacteria under the cyanobacteria rubisco promoter. | <p>Dry land growth cyanobacteria have strongly abilities to survive in desert. Therefore, it can be used as an ideal bioreactor in for producing cellulose which is a good material for water retention. We cloned and over-expressed seven bacterial genes (BCS genes) which are responsible for cellulose synthesis into cyanobacteria under the cyanobacteria rubisco promoter. | ||
The genetic transferred cyanobacteria was shown in Figure 1.In addition, the production of cellulose in the transgenic cyanobacteria was analyzed(Figure 2). | The genetic transferred cyanobacteria was shown in Figure 1.In addition, the production of cellulose in the transgenic cyanobacteria was analyzed(Figure 2). | ||
| Line 461: | Line 461: | ||
<p style="font-size:16px; text-align: center;">Figure 2 The measurement of cellulose content in transgenic cyanobacteria with bcs gene.</p></div> | <p style="font-size:16px; text-align: center;">Figure 2 The measurement of cellulose content in transgenic cyanobacteria with bcs gene.</p></div> | ||
| − | <div class="DBoard" id="title" style="height:40px;width: | + | <div class="DBoard" id="title" style="height:40px;width:600px;font-size:20px;line-height:40px;text-align:center;"> |
2.Mathematical Model of Biological Intrinsic Regulation System (HAWNA) | 2.Mathematical Model of Biological Intrinsic Regulation System (HAWNA) | ||
</div> | </div> | ||
| Line 473: | Line 473: | ||
<div class="DBoard" id="article1" style="height:260px;"> | <div class="DBoard" id="article1" style="height:260px;"> | ||
<p>Basing on the metabolic pathway of bacterial cellulose, knocking out gene slr0897 is likely to improve the production of bacterial cellulose. However, it could also effect the entire metabolic network since it is a key gene involved in many metabolic pathways. By using computer modeling, we are able to predict genes that may positively regulate slr0897 but have little effects on the growth of Synechocystis sp. We have identified a gene named PCC6803. Knocking it out may increase the yield of bacterial cellulose. </p></div> | <p>Basing on the metabolic pathway of bacterial cellulose, knocking out gene slr0897 is likely to improve the production of bacterial cellulose. However, it could also effect the entire metabolic network since it is a key gene involved in many metabolic pathways. By using computer modeling, we are able to predict genes that may positively regulate slr0897 but have little effects on the growth of Synechocystis sp. We have identified a gene named PCC6803. Knocking it out may increase the yield of bacterial cellulose. </p></div> | ||
| − | <div class="DBoard" id="article2" align="center" style="height: | + | <div class="DBoard" id="article2" align="center" style="height:935px;"> |
| − | <img src="https://static.igem.org/mediawiki/2018/b/b1/T--SCAU-China--model15.jpg" width=" | + | <img src="https://static.igem.org/mediawiki/2018/e/e5/T--SCAU-CHINA--demonstrate12.jpg" width="430px" height="530px" /><br><br><br> |
| − | <p style="font-size:16px; text-align: center;">Figure 4 | + | <img src="https://static.igem.org/mediawiki/2018/b/b1/T--SCAU-China--model15.jpg" width="800px" height="300px" /> |
| + | <p style="font-size:16px; text-align: center;">Figure 4 the illustration of Causal network analysis and the heat map of non-target pathway genes which positive correlated with slr0897 </p></div> | ||
<div class="DBoard" id="title" style="height:40px;width:400px;font-size:20px;line-height:40px;text-align:center;"> | <div class="DBoard" id="title" style="height:40px;width:400px;font-size:20px;line-height:40px;text-align:center;"> | ||
| Line 490: | Line 491: | ||
<p><b>Experimental Result</b></p></div> | <p><b>Experimental Result</b></p></div> | ||
<div class="DBoard" id="article2" align="center"> | <div class="DBoard" id="article2" align="center"> | ||
| − | <img src="https://static.igem.org/mediawiki/2018/8/8c/T--SCAU-CHINA--demonstrate6.PNG" width=" | + | <img src="https://static.igem.org/mediawiki/2018/8/8c/T--SCAU-CHINA--demonstrate6.PNG" width="360px" height="430px" /> |
<p style="font-size:16px; text-align: center;">Figure 6: The GFP intensity measurement result demonstrating the promoter activity which is consistent to our prediction. </p> | <p style="font-size:16px; text-align: center;">Figure 6: The GFP intensity measurement result demonstrating the promoter activity which is consistent to our prediction. </p> | ||
</div> | </div> | ||
| Line 504: | Line 505: | ||
</p></div> | </p></div> | ||
<div class="DBoard" id="article2" align="center"> | <div class="DBoard" id="article2" align="center"> | ||
| − | <img src="https://static.igem.org/mediawiki/2018/a/a8/T--SCAU-China--mc-1.jpg" width=" | + | <img src="https://static.igem.org/mediawiki/2018/a/a8/T--SCAU-China--mc-1.jpg" width="380px" height="500px" /> |
<p style="font-size:16px; text-align: center;">Figure7. The flowchart of our cell growth characterized model </p></div> | <p style="font-size:16px; text-align: center;">Figure7. The flowchart of our cell growth characterized model </p></div> | ||
<div class="DBoard" id="article1" style="height:160px;"> | <div class="DBoard" id="article1" style="height:160px;"> | ||
Revision as of 08:07, 17 October 2018
Dry land growth cyanobacteria have strongly abilities to survive in desert. Therefore, it can be used as an ideal bioreactor in for producing cellulose which is a good material for water retention. We cloned and over-expressed seven bacterial genes (BCS genes) which are responsible for cellulose synthesis into cyanobacteria under the cyanobacteria rubisco promoter. The genetic transferred cyanobacteria was shown in Figure 1.In addition, the production of cellulose in the transgenic cyanobacteria was analyzed(Figure 2). We have successfully over-produced the cellulose in the transgenic cyanobacteria. The glucose (bacterial cellulose digestion produce) content of the transgenic is much higher that the WT (Figure 2). There was significant differences between and wild-type. Unfortunately, the growth of the transgenic stain was much slower than the WT (Figure 1).
These results indicate that the bacterial cellulose can be over-expressed and accumulated in the cyanobacteria. We need to reduce the negative impact of gene over-expression on the normal metabolism in cyanobacteria and meanwhile further increase the production levels of bacterial cellulose.

Figure 1 Transferred cyanobacteria (the left) and the wail type (the right).The transferred cyanobacteria is inhibited in the growth (in yellow color) but not in cellulose production.
Here is the measurement of cyanobacteria glucose (3 repeats). The same of amount of transgenic cyanobacteria with bcsZH-ABCD-bglX genes and wild-type were treated with lysozyme to break the cells. Due to lacking a direct way to measure the content of cellu;ose in bacterial cell wall. Therefore, glucose can be used as an alternative parameter for measuring the content of cellulose since it can be digested into glucose by cellulose.
The differences between the red column and blue column indicated that the content of bacteria cellulose.

Figure 2 The measurement of cellulose content in transgenic cyanobacteria with bcs gene.
To better solve the problems that we encountered above during over-production of cellulose in cyanobacteria, we took advantage of applying computer modeling and system biology into our synthetic biology experiments. We aimed to manipulate the BCS genes expression network and underpin the key factors that directly regulate the cellulose gene expression. However, gene regulating network in living organisms is extremely complicated, and any modification on genetic node may lead to the lateration of the entire metablic network. Therefore, Iwe carefully developed an system biology method termed as HAWNA (Hierarchical And Weighted Network Analysis). It combined with gene and prorein expression profiles to underpin the key regulatory genes that contribute directly for the metabolic network and reduce the noise of gene expression profile significantly.

Figure3 The structure of HAWNA (Need to insert a new image above!)
Basing on the metabolic pathway of bacterial cellulose, knocking out gene slr0897 is likely to improve the production of bacterial cellulose. However, it could also effect the entire metabolic network since it is a key gene involved in many metabolic pathways. By using computer modeling, we are able to predict genes that may positively regulate slr0897 but have little effects on the growth of Synechocystis sp. We have identified a gene named PCC6803. Knocking it out may increase the yield of bacterial cellulose.

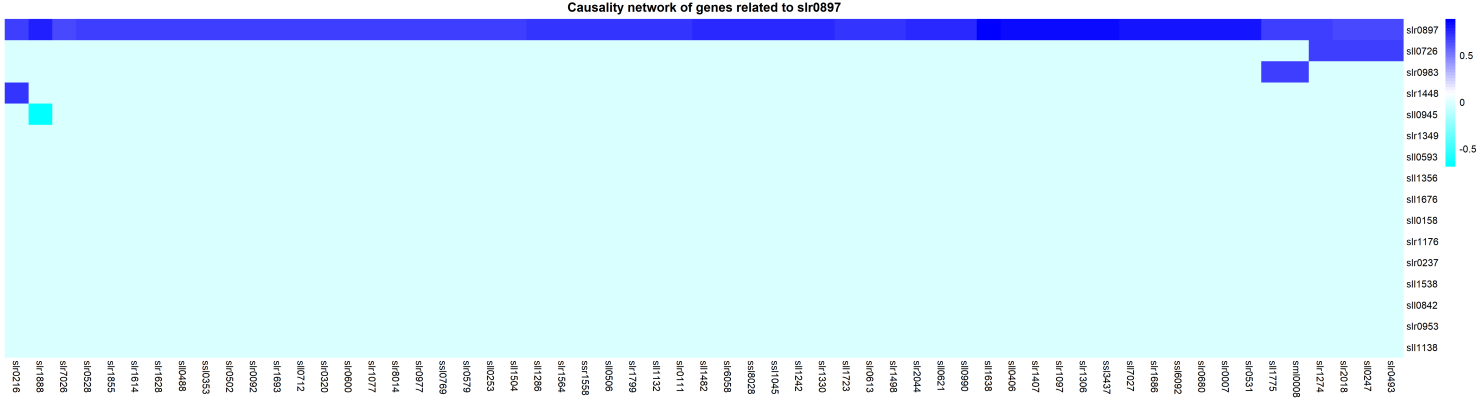
Figure 4 the illustration of Causal network analysis and the heat map of non-target pathway genes which positive correlated with slr0897
Our team proposes a new computational method to predict the intensity of promoters in E.coil, and efficiently identify their most importance sequencess or units which act as the core element to determine the activities of promoters. We used the sequence information to predict promoter activity. our training set sequence includes both original promoters from E.coil and recomposed promoters which are caused by the infection of lambda bacteriophage. In addition, we combined a new feature encoding method with machine learning algorithm to predict the promoter strength and consensus unit or core site may have strong correlation to the activities of the promoter.

Figure 5 The flowchart of feature encoding. We summarized the unit frequency and encode each sequence site as a numeric format. Then concatenate above two vectors into a 132-dimensional feature vector to predict promoter activity
Using our model, we predicted and measured a wide range of E.coil promoter activities based on the Ecoil promoter database ‘PromEC’ which includes 471 promoter sequences [4](see predict_promEC.txt). According to our prediction, we random selected three promoters: rplj (predict value:1.723286152), dapA (predict value:1.235077024) and caiF (predict value:0.68780911) to make chimeric fusion proteins with green fluorescent protein (GFP) test the activities of these three promoter to test and verify our prediction.
Experimental Result

Figure 6: The GFP intensity measurement result demonstrating the promoter activity which is consistent to our prediction.
To improve the production of bacterial cellulose in cyanobacteria, it is necessary to increase our cultured cell biomass. In vivo, we have intergrated the existing literature data base to build an interactome to help us design a new scheme to boost the cellulose production via metabolic engineering. In vitro, based on measured data tracing Synechocystis growth, we constructed a two layers of model to characterize the main parameters influencing the behavior of our system.
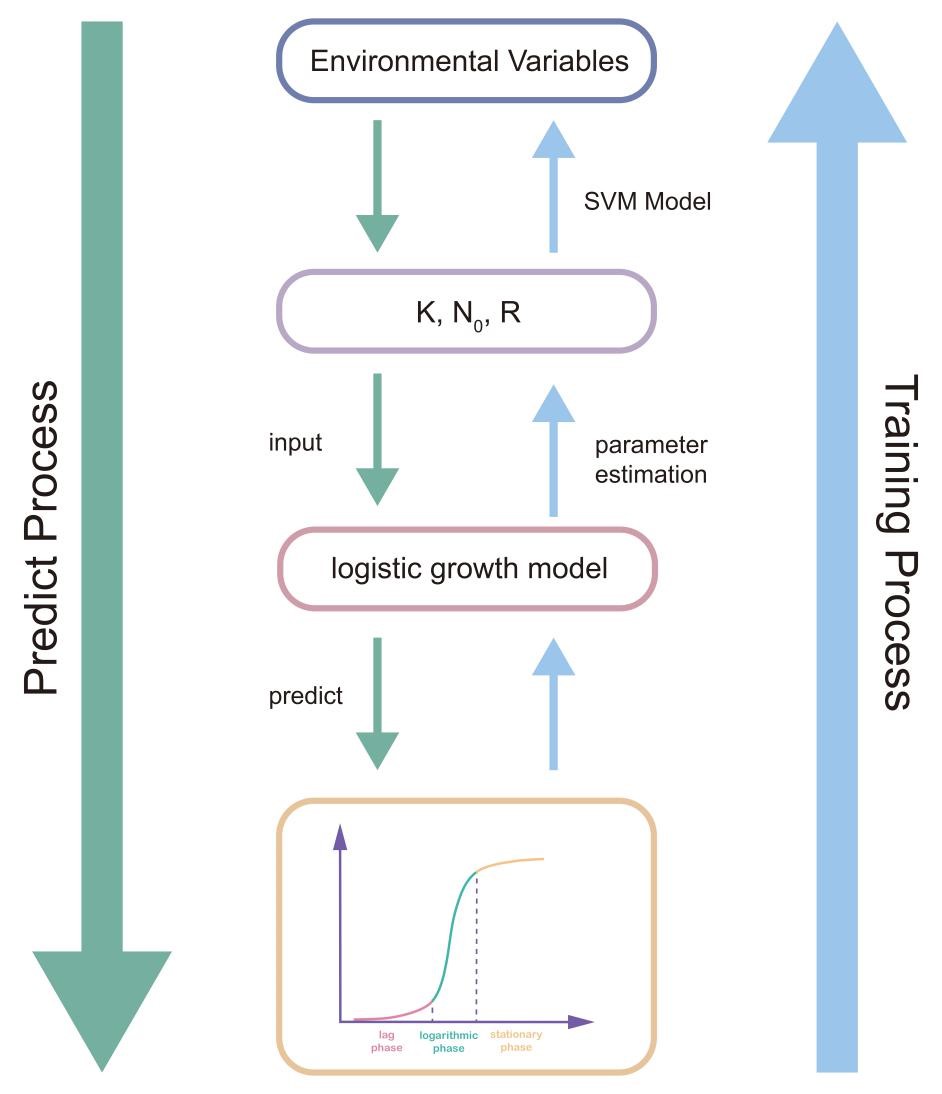
Figure7. The flowchart of our cell growth characterized model
We simulated about 1875 combinations, and found the following conditions were critical for the best K and R value compare with other schemes. Besides, we also simulated its growth curve by comparion with our measurement of cynobacteria growth curve. It showed that our cultured conditions can be optimized.

Figure8 The best K and R value for the optimization of cell culture.
We verified through experiments that this Synergistic Recombination works well in Synechocystis and Microcoleus, with PgroESL, Prbc, Trbc and slr0168. It is conceptual and in dynamic change, expected to be applied to a wider range of fields.


Figure 9 PCR amplifying of homologous genes slr0168 in Synechocystis and Microcoleus
And, we designed a new Cas9 Knocking out Kit (Type II CRISPR system) based on the Synergistic Recombination kit experiment to knock out Slr0163/Slr0977/Slr0897/Slr1278.

Figure 10 Colony PCR for the construction of Cas 9 knocking out kit.
Taken together, we achieved bacterial cellulose expression in cyanobacteriawhich can be further used as a brand new strategy for desertification controllfrom synthetic biology aspectof view. In order to increase the cellulose production and optimize the growth conditions of the transgenic cyanobacteria, we computer model the metabolic pathway of cellulose synthesis, the culture condition as well as prediction of gene promoter intensity of our genetic transferred genes. Finally, we designed two useful kits for the transformation between different species cyanobacteria. Cyanobacteria can be regarded a ideal bio-reactor for water retention in desert fordesertification control.
School's name:SCAU
Member's name:SCAU
Designed by:SCAU